Gli antipiretici per i bambini sono prescritti da un pediatra. Ma ci sono situazioni di emergenza per la febbre quando il bambino ha bisogno di ricevere immediatamente la medicina. Quindi i genitori si assumono la responsabilità e usano farmaci antipiretici. Cosa è permesso dare ai neonati? Come abbassare la temperatura nei bambini più grandi? Quali farmaci sono i più sicuri?
Assone (greco ἀξον - asse) - neurite, cilindro assiale, processo di una cellula nervosa, lungo il quale impulsi nervosi andare dal corpo cellulare (soma) agli organi innervati e ad altre cellule nervose.
Un neurone è costituito da un assone, un corpo e diversi dendriti, a seconda del numero di cui le cellule nervose sono suddivise in unipolari, bipolari, multipolari. La trasmissione dell'impulso nervoso avviene dai dendriti (o dal corpo cellulare) all'assone, quindi il potenziale d'azione generato dal segmento iniziale dell'assone viene ritrasmesso ai dendriti. Se un assone nel tessuto nervoso si connette con il corpo della cellula nervosa successiva, tale contatto è chiamato asso-somatico, con dendriti - asso-dendritico, con un altro assone - asso-assonale (un raro tipo di connessione, che si trova nel centro sistema nervoso).
Alla giunzione dell'assone con il corpo del neurone, le più grandi cellule piramidali del quinto strato della corteccia hanno un tumulo di assoni. In precedenza, si presumeva che qui avvenisse la conversione del potenziale postsinaptico del neurone in impulsi nervosi, ma i dati sperimentali non lo confermavano. La registrazione dei potenziali elettrici ha rivelato che l'impulso nervoso è generato nell'assone stesso, vale a dire nel segmento iniziale a una distanza di ~50 µm dal corpo del neurone. Per generare un potenziale d'azione nel segmento iniziale dell'assone è necessaria una maggiore concentrazione di canali del sodio (fino a cento volte rispetto al corpo del neurone).
La nutrizione e la crescita dell'assone dipendono dal corpo del neurone: quando l'assone viene tagliato, la sua parte periferica muore, mentre la parte centrale rimane vitale. Con un diametro di diversi micron, la lunghezza dell'assone può raggiungere 1 metro o più in animali di grandi dimensioni (ad esempio, assoni provenienti dai neuroni del midollo spinale agli arti). Molti animali (calamari, pesci, anellidi, foronidi, crostacei) hanno assoni giganti spessi centinaia di micron (fino a 2-3 mm nei calamari). Tipicamente, questi assoni sono responsabili della conduzione dei segnali ai muscoli. fornendo una "reazione di fuga" (attirare in un visone, nuotare velocemente, ecc.). Ceteris paribus, con un aumento del diametro dell'assone, aumenta la velocità di conduzione degli impulsi nervosi lungo di esso.
Nel protoplasma dell'assone - axoplasm - ci sono le fibre più sottili - neurofibrille, oltre a microtubuli, mitocondri e un reticolo endoplasmatico agranulare (liscio). A seconda che gli assoni siano ricoperti da una guaina mielinica (polpa) o privi di essa, formano fibre nervose polpose o amielinizzate.
La guaina mielinica degli assoni si trova solo nei vertebrati. È formato da speciali cellule di Schwann "avvolte" sull'assone, tra le quali vi sono aree libere dalla guaina mielinica - le intercettazioni di Ranvier. Solo alle intercettazioni sono presenti i canali del sodio voltaggio-dipendenti e ricompare il potenziale d'azione. In questo caso, l'impulso nervoso si propaga lungo le fibre mielinizzate a gradini, il che aumenta più volte la velocità della sua propagazione.
Le sezioni terminali dell'assone - terminali - si ramificano e contattano altre cellule nervose, muscolari o ghiandolari. Alla fine dell'assone c'è una terminazione sinaptica - la sezione terminale del terminale a contatto con la cellula bersaglio. Insieme alla membrana postsinaptica della cellula bersaglio, la terminazione sinaptica forma una sinapsi. L'eccitazione viene trasmessa attraverso le sinapsi.
Il trasporto dell'assone è il movimento lungo l'assone di una cellula nervosa di vario materiale biologico.
I processi assonali dei neuroni sono responsabili della trasmissione del potenziale d'azione dal corpo del neurone alla sinapsi. Inoltre, l'assone è un percorso lungo il quale vengono trasportati i materiali biologici necessari tra il corpo del neurone e la sinapsi, necessari per il funzionamento della cellula nervosa. Organelli di membrana (mitocondri), varie vescicole, molecole di segnalazione, fattori di crescita, complessi proteici, componenti citoscheletrici e persino canali Na + e K + vengono trasportati lungo l'assone dall'area di sintesi nel corpo del neurone. I punti finali di questo trasporto sono determinate aree dell'assone e della placca sinaptica. A loro volta, i segnali neurotrofici vengono trasportati dalla sinapsi al corpo cellulare. Questo agisce come un feedback che riporta lo stato di innervazione del bersaglio.La lunghezza dell'assone del sistema nervoso periferico umano può superare 1 m, e può essere maggiore nei grandi animali. Lo spessore di un grande motoneurone umano è di 15 micron, che, con una lunghezza di 1 m, dà un volume di ~ 0,2 mm³, che è quasi 10.000 volte più volume cellule epatiche. Ciò rende i neuroni dipendenti dal trasporto fisico efficiente e coordinato di sostanze e organelli lungo gli assoni.
Le lunghezze e i diametri degli assoni, così come la quantità di materiale trasportato lungo di essi, indicano certamente la possibilità di guasti ed errori nel sistema di trasporto. Molte malattie neurodegenerative sono direttamente correlate a disturbi nel funzionamento di questo sistema.
Semplificato, il trasporto di assoni può essere rappresentato come un sistema costituito da più elementi. Comprende il carico, le proteine motrici che effettuano il trasporto, i filamenti del citoscheletro o "rotaie" lungo le quali i "motori" sono in grado di muoversi. Sono inoltre necessarie proteine linker che collegano le proteine motrici al loro carico o ad altre strutture cellulari e molecole ausiliarie che avviano e regolano il trasporto.
Le proteine del citoscheletro vengono rilasciate dal corpo cellulare, muovendosi lungo l'assone a una velocità da 1 a 5 mm al giorno. Questo è un trasporto lento di assoni (un trasporto simile è presente anche nei dendriti). Anche molti enzimi e altre proteine citosoliche sono trasportate da questo tipo di trasporto.I materiali non citosolici necessari alla sinapsi, come le proteine secrete e le molecole legate alla membrana, si muovono lungo l'assone a una velocità molto maggiore. Queste sostanze vengono trasportate dal loro sito di sintesi, il reticolo endoplasmatico, all'apparato di Golgi, che spesso si trova alla base dell'assone. Quindi queste molecole, impacchettate in vescicole di membrana, vengono trasportate lungo i binari dei microtubuli mediante un rapido trasporto assonale a una velocità fino a 400 mm al giorno. Pertanto, i mitocondri, varie proteine, inclusi i neuropeptidi (neurotrasmettitori di natura peptidica), i neurotrasmettitori non peptidici vengono trasportati lungo l'assone.Il trasporto di materiali dal corpo del neurone alla sinapsi è chiamato anterogrado e nella direzione opposta - retrogrado. Il trasporto lungo l'assone su lunghe distanze avviene con la partecipazione dei microtubuli. I microtubuli nell'assone hanno la loro polarità intrinseca e sono orientati con un'estremità in rapida crescita (più) verso la sinapsi e un'estremità in crescita lenta (meno) verso il corpo del neurone. Le proteine motorie di trasporto degli assoni appartengono alle superfamiglie chinesina e dineina.Le chinesine sono principalmente proteine motorie più-terminali che trasportano merci come i precursori delle vescicole sinaptiche e gli organelli di membrana. Questo trasporto va verso la sinapsi (anterograda). Le dineine citoplasmatiche sono proteine motorie meno terminali che trasportano segnali neurotrofici, endosomi e altro carico retrogrado al corpo del neurone. Il trasporto retrogrado non è esclusivo delle dineine: sono state trovate diverse kinesine che si muovono in direzione retrograda.
11.Fibre mielinizzate e non mielinizzate. processo di mielinizzazione. Il dolore nervoso contiene fibre mielinizzate e non mielinizzate o debolmente mielinizzate. La composizione cellulare degli spazi endoneurali riflette il livello di mielinizzazione. Normalmente, il 90% dei nuclei cellulari trovati in questo spazio appartiene alle cellule di Schwann (lemmociti) e il resto appartiene ai fibroblasti e all'endotelio capillare. All'80%, le cellule di Schwann circondano gli assoni non mielinizzati; accanto alle fibre mielinizzate, il loro numero si riduce di 4 volte. Le fibre mielinizzate di grande diametro conducono impulsi molto di più ritmo veloce piuttosto che debolmente mielinizzato o non mielinizzato. Esistono tre classi di fibre: A, B e C. Fibre A - fibre nervose mielinizzate afferenti e afferenti somatiche, fibre B - fibre vegetative pregangliari mielinizzate, fibre C - fibre autonomiche e sensoriali non mielinizzate. La mielina copre la guaina dei tronchi nervosi e garantisce una trasmissione più efficiente dell'impulso nervoso. Il processo è chiamato mielinizzazione perché il risultato è una guaina di mielina, che è circa 2/3 di grasso ed è un buon isolante elettrico. I ricercatori attribuiscono grande importanza al processo di mielinizzazione nello sviluppo del cervello. È noto che circa i 2/3 delle fibre cerebrali di un neonato sono mielinizzate. A circa 12 anni, la fase successiva della mielinizzazione è completata. Ciò corrisponde al fatto che il bambino sta già formando la funzione dell'attenzione, ha abbastanza bene il controllo di se stesso. Tuttavia, il processo completo di mielinizzazione termina solo alla fine della pubertà. Pertanto, il processo di mielinizzazione è un indicatore della maturazione di una serie di funzioni mentali. Si scopre che le fibre mielinizzate conducono l'eccitazione centinaia di volte più velocemente di quelle non mielinizzate, cioè le reti neurali del nostro cervello possono funzionare più velocemente e quindi in modo più efficiente.
12.Connessioni interneuronali. Sinapsi, loro struttura e funzioni. Nelle fasi successive della filogenesi, e soprattutto nell'uomo, la connessione tra le cellule nervose viene effettuata da formazioni speciali: le sinapsi. La sinapsi è costituita da tre elementi principali: la membrana presinaptica, la fessura sinaptica e la membrana postsinaptica. La membrana presinaptica è un apparato neurosecretorio in cui viene sintetizzato e rilasciato un mediatore, che ha un effetto inibitorio o eccitatorio sulla membrana postsinaptica della cellula innervata. La membrana postsinaptica ha una sensibilità selettiva a un agente chimico - un mediatore ed è praticamente insensibile a uno stimolo elettrico. La presenza di sinapsi determina la conduzione unilaterale di un impulso nervoso (la trasmissione inversa dell'eccitazione dalla membrana postsinaptica a quella presinaptica è impossibile), poiché l'eccitazione può diffondersi in entrambe le direzioni dallo stimolo nelle fibre nervose. Allo stesso tempo, la velocità di conduzione rallenta nella sinapsi. La durata del ritardo sinaptico varia notevolmente a seconda dello scopo funzionale della sinapsi ed è di 0,2-0,5 ms nelle sinapsi interneuronali e neuromuscolari, mentre nelle terminazioni nervose della muscolatura liscia raggiunge i 5-10 ms.
13.Tipi di sinapsi (chimiche ed elettriche). Meccanismo di trasmissione sinaptica. Il mediatore situato nelle vescicole viene rilasciato nella fessura sinaptica per esocitosi (le vescicole si avvicinano alla membrana, si fondono con essa e si rompono, rilasciando il mediatore). Il suo rilascio avviene in piccole porzioni - quanti. Ogni quanto contiene da 1.000 a 10.000 molecole di neurotrasmettitore. Un piccolo numero di quanti esce dal finale e sono a riposo. Quando l'impulso nervoso, ad es. AP raggiunge l'estremità presinaptica, si verifica la depolarizzazione della sua membrana presinaptica. aprila canali del calcio e gli ioni calcio entrano nella placca sinaptica. Inizia il rilascio di un gran numero di quanti di neurotrasmettitori. Le molecole trasmittenti diffondono attraverso la fessura sinaptica alla membrana postsinaptica e interagiscono con i suoi chemocettori. Come risultato della formazione di complessi mediatore-recettore, inizia la sintesi dei cosiddetti messaggeri secondari nella membrana subsinaptica. In particolare cAMP. Questi mediatori attivano i canali ionici nella membrana postsinaptica. Pertanto, tali canali sono chiamati chemodipendenti o recettoriali. Quelli. si aprono sotto l'azione della PAS sui chemocettori. Come risultato dell'apertura dei canali, il potenziale della membrana subsinaptica cambia. Questo cambiamento è chiamato potenziale postsinaptico. sinapsi elettriche. è una formazione simile a una fessura (dimensioni della fessura fino a 2 nm) con ponti ionici-canali tra due cellule a contatto. Gli anelli di corrente, in particolare in presenza di un potenziale d'azione (AP), saltano quasi senza ostacoli attraverso un tale contatto simile a una fessura ed eccitano, cioè indurre la generazione di AP della seconda cella. In generale, tali sinapsi (sono chiamate efapsi) forniscono una trasmissione molto rapida dell'eccitazione. Ma allo stesso tempo, la conduzione unilaterale non può essere assicurata con l'aiuto di queste sinapsi, poiché la maggior parte di queste sinapsi ha una conduzione bidirezionale. Inoltre, non possono essere utilizzati per forzare una cellula effettrice (una cellula controllata attraverso una data sinapsi) a inibire la sua attività. Un analogo della sinapsi elettrica nei muscoli lisci e nel muscolo cardiaco sono le giunzioni di tipo nexus. sinapsi chimiche. Per struttura, le sinapsi chimiche sono terminazioni di assoni (sinapsi terminali) o la sua parte varicosa (sinapsi di passaggio), che è piena di una sostanza chimica - un mediatore. Nella sinapsi si distingue un elemento presinaptico, che è limitato dalla membrana presinaptica, un elemento postsinaptico, che è limitato dalla membrana postsinaptica, nonché una regione extrasinaptica e una fessura sinaptica, la cui dimensione media è di 50 nm. C'è un'ampia varietà nei nomi delle sinapsi in letteratura. Ad esempio, una placca sinaptica è una sinapsi tra neuroni, una placca terminale è una membrana postsinaptica di una sinapsi mioneurale, una placca motoria è una terminazione presinaptica di un assone su una fibra muscolare.
Nel trasferimento di informazioni a sistema nervoso coinvolti, i loro processi e le sinapsi. All'interno delle sinapsi, le informazioni vengono trasferite da un neurone all'altro sostanze chimiche- mediatori.
Dendriti e assoni dei neuroni. Le cellule nervose sono bipolari, il che significa la trasmissione di informazioni in una sola direzione: un'estremità della cellula riceve informazioni e l'altra le invia ad altri neuroni. Il corpo del neurone è coperto enorme quantità processi densamente ramificati - dendriti, progettati per ricevere informazioni.
Numero dendriti e la natura della loro ramificazione in diversi neuroni può variare in modo significativo. Il processo di una cellula nervosa che trasmette ulteriormente gli impulsi nervosi è chiamato assone; la lunghezza dell'assone nell'uomo può raggiungere 1 M. "Cilindro assiale" è un nome descrittivo obsoleto e ormai poco utilizzato per l'assone, che ne enfatizzava la forma: un cilindro allungato.
assone- una struttura unica, perché possono esserci molti dendriti, ma un assone è sempre uno. L'estremità distale dell'assone termina con molti rami terminali (terminali e naleys), ognuno dei quali si espande come una gemma (estensione terminale, terminazione sinaptica, placca sinaptica); questa formazione entra in contatto con la cellula successiva.
Separato menzioni meritano lunghi processi periferici di neuroni pseudo-unipolari nei nodi spinali, che portano informazioni al sistema nervoso centrale sulla sensibilità tattile, al dolore e alla temperatura dalla superficie del corpo. Il fatto è che venivano anche chiamati assoni per la loro somiglianza puramente strutturale, senza tener conto del fatto che questi processi ricevono informazioni.
Trofico, cioè nutriente, centro neurone serve il suo corpo (soma, o perikaryon), che contiene il nucleo e vari organelli.
Trasporto assonale (assone) dei neuroni. Nel corpo vengono sintetizzati mediatori o enzimi che catalizzano la loro biosintesi, che poi si muovono lungo i microtubuli degli assoni fino all'estremità dell'assone - questo processo è chiamato trasporto diretto dell'assone.
All'interno delle estensioni finali assone sotto forma di vescicole (vescicole sinaptiche) vengono immagazzinate molecole di mediatori, molte delle quali si accumulano in ciascuna terminazione sinaptica. Tuttavia, il trasporto dell'assone viene effettuato in entrambe le direzioni, non solo dal corpo del neurone all'estremità dell'assone (trasporto diretto, anterogrado), ma anche all'indietro (trasporto inverso, retrogrado).
Velocità trasporto rapido degli assoni(attraverso i microtubuli con il coinvolgimento di energia) è di 200-400 mm/giorno e la velocità della corrente axoplasmica lenta non supera 1-5 mm/giorno. Il trasporto degli assoni è preso come base per il metodo dei segni anterogradi e retrogradi, attraverso i quali viene tracciato il corso dei processi neuronali.
Myelination di assoni di neuroni. Gli assoni dei neuroni sono avvolti in una guaina di mielina. Nel sistema nervoso centrale, la guaina mielinica (polpa) è formata da oligodendrociti (un tipo speciale di cellule gliali) e nel sistema nervoso periferico da cellule di Schwann o cellule di Schwann. La membrana di un oligodendrocita o di un lemmocita estende un processo verso l'assone, che, espandendosi come una frittella, avvolge ripetutamente l'assone e fornisce il suo isolamento elettrico.
Un mucchio di oligodendrociti o le cellule di Schwann possono avvolgere sequenzialmente un assone. Due cellule adiacenti e uno spazio assonale nudo tra di loro (nodo nodale di Ranvier) è considerato un segmento della guaina mielinica. Poiché la mielina svolge il ruolo di isolante, il potenziale d'azione (AP) provoca la depolarizzazione esclusivamente ai nodi di Ranvier, motivo per cui eccitazione nervosa come se "saltasse" lungo l'assone da un'intercettazione all'altra. Questo processo è chiamato conduzione saltatoria. Le fibre mielinizzate più spesse con un ampio intervallo tra i nodi di Ranvier conducono l'eccitazione nervosa più velocemente di altre.
Per assoni, povero mielina, l'eccitazione, al contrario, si diffonde relativamente lentamente, poiché è costretta a "diffondersi" su tutta la membrana della fibra. Gli assoni mielinizzati più sottili occupano una posizione intermedia tra di loro. Pertanto, gli assoni sono divisi in mielinizzati spessi (A), mielinizzati sottili (B) e non mielinizzati (C). Il diametro delle fibre spesse di mielina di tipo A è di 3-20 μm, la velocità di eccitazione attraverso queste fibre raggiunge i 120 m/s. Il diametro delle sottili fibre mieliniche di tipo B è di 3 µm e la velocità di conduzione è di 15 m/s; la velocità di conduzione lungo le fibre non mielinizzate di tipo C non supera i 2 m/s.
Il sistema nervoso umano contiene almeno un trilione (10 12) di cellule nervose, circa 10 13 cellule gliali e non meno (> 10 13) sinapsi. Il numero di tipi di cellule è sconosciuto (almeno 100). Questo insieme forma una struttura spaziale complessa: un'unica rete con numerose connessioni sia a livello di una singola cellula che di insiemi cellulari (CNS).
TESSUTO NERVOSO
Elementi istologici tessuto nervoso(neuroni e gliociti) e gli organi sensoriali si sviluppano da derivati del neuroectoderma, vale a dire dal tubo neurale, dalla cresta neurale e dai placodi neurogenici. La neurulazione, durante la quale si forma il neuroectoderma, è discussa nel Capitolo 3 (vedi Figura 3-13).
tubo neurale contiene cellule neurali staminali(sono anche matrici o ventricolari) - la fonte di quasi tutte le cellule del SNC. Le cellule ventricolari proliferano e danno origine a neuroblasti e glioblasti cellule progenitrici neuroni e gliociti. Rimane parte delle cellule ventricolari sul posto questo è il futuro ependima. Cresta neurale. Se il tubo neurale funge da fonte per lo sviluppo del sistema nervoso centrale, allora gli elementi nervosi della periferia e una serie di altre strutture importanti (vedi Tabelle 3-3) hanno origine dalla cresta neurale. placodi neurogeni. Formano neuroni del rivestimento olfattivo, gangli vestibolari e uditivi, nonché neuroni sensoriali dei gangli genicolato, pietroso, nodulare e trigemino. nervi cranici.
Neuroblasti- cellule con un grande nucleo arrotondato, nucleolo denso e citoplasma pallido - danno origine ai neuroni. I neuroni appartengono a una popolazione statica. In nessun caso lo sono in vivo incapace di proliferare e rinnovarsi. L'eccezione sono i neuroni olfattivi del rivestimento epiteliale del nasale
passaggi di uscita, così come alcuni neuroni dell'ippocampo e del bulbo olfattivo.
Glioblasti- precursori della macroglia (astrociti e oligodendrociti). Tutti i tipi di macroglia sono in grado di proliferare.
Microglia. Le cellule microgliali sono classificate come fagociti mononucleari.
Neuroni
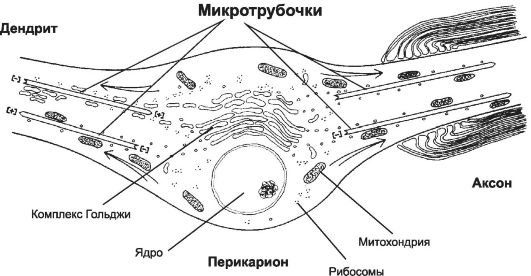
I neuroni sono i principali tipi di cellule del tessuto nervoso. Queste cellule eccitabili conducono segnali elettrici e forniscono la capacità del cervello di elaborare le informazioni. Il pericarion (il corpo di un neurone), i processi che si estendono da esso (assone e dendriti) sono le parti standard dei neuroni (Fig. 8-13).
Pericarion contiene un nucleo, complesso di Golgi, reticolo endoplasmatico granulare, mitocondri, lisosomi, elementi del citoscheletro.
Nucleo Il neurone ha cromatina finemente dispersa e un nucleolo. A causa del diametro relativamente grande, il nucleo appare otticamente vuoto al microscopio ottico. Il nucleolo è grande e nettamente basofilo.
Riso. 8-13. Neurone multipolare. Il corpo cellulare (pericarion) contiene il nucleo. I processi derivano dal pericarion. Uno di questi è un assone, tutti gli altri sono dendriti.
Complesso Golgi è ben sviluppato. La sua particolarità è la sua posizione tra il nucleo e l'origine dell'assone, che riflette il potente trasporto di proteine sintetizzate nel reticolo endoplasmatico granulare del pericarion nell'assone.
collinetta assonica- una regione del pericarion priva di reticolo endoplasmatico granulare e ribosomi, contenente molti microtubuli e neurofilamenti, questo è il luogo in cui inizia l'assone e si genera il potenziale d'azione.
Reticolo endoplasmatico granulare. Un reticolo endoplasmatico granulare si sviluppa nel pericarion e nei dendriti. Corrisponde a grumi di sostanza cromatofila (sostanza di Nissl).
Mitocondri numerose. Fabbisogno energetico significativo cellule nervose fornisce un metabolismo prevalentemente aerobico, motivo per cui i neuroni sono estremamente sensibili all'ipossia.
Pigmenti. Nei neuroni (soprattutto con l'età), la lipofuscina si accumula. I neuroni di alcuni nuclei cerebrali normalmente contengono altri pigmenti, motivo per cui queste formazioni hanno preso il loro nome (substantia nigra, locus coeruleus).
germogli, che si estende dal pericarion - l'assone e i dendriti (Fig. 8-13).
assone(neurite) - un lungo processo, di regola, che non si ramifica per tutta la sua lunghezza, ma forma rami terminali contenenti vescicole sinaptiche. Il volume di un assone può raggiungere il 99% del volume totale di un neurone. La lunghezza dell'assone può essere piuttosto significativa, fino a decine di centimetri.
Dendriti- processi di ramificazione che terminano vicino al corpo del neurone. I recettori postsinaptici sono incorporati nel plasmolemma, i dendriti conducono l'eccitazione al pericarion. Le aree prossimali dei dendriti sono una continuazione del pericarion. Pertanto, contengono ribosomi, componenti del reticolo endoplasmatico granulare e liscio, elementi del complesso del Golgi. I dendriti formano integralmente fino al 95% dell'intera superficie del recettore (campo recettivo) del neurone.
citoscheletro I neuroni sono costituiti da microtubuli, filamenti intermedi (neurofilamenti) e microfilamenti.
microtubuli- gli elementi più grandi del citoscheletro, il loro diametro è di 24 nm. Sono associati all'intracellulare, incluso trasporto di assoni. Varie sostanze (proteine, neurotrasmettitori, ecc.), Organelli (mitocondri, elementi citoscheletrici, vescicole, ecc.) Si spostano dal pericarion lungo i processi. I microtubuli nel pericarion e nei dendriti (a differenza dell'assone) non hanno un orientamento direzionale. La maggior parte dei microtubuli dell'assone (+)-estremità sono diretti al terminale e (-)-estremità - al pericarion (Fig. 8-14). La natura dell'orientamento dei microtubuli ha importanza per la distribuzione lungo i processi di vari organelli. I mitocondri e le vescicole secretorie si spostano all'estremità (+) e i ribosomi, i corpi multivescicolari, gli elementi del complesso di Golgi si spostano all'estremità (-).
 Riso. 8-14. Orientamento dei microtubuli nei processi neuronali. L'assone differisce dai dendriti nell'orientamento polare dei microtubuli. In esso, i microtubuli con le loro (+) - estremità sono dirette al terminale e (-) - terminano - al pericarion.
Riso. 8-14. Orientamento dei microtubuli nei processi neuronali. L'assone differisce dai dendriti nell'orientamento polare dei microtubuli. In esso, i microtubuli con le loro (+) - estremità sono dirette al terminale e (-) - terminano - al pericarion.
Classificazione dei neuroni. I neuroni differiscono per le dimensioni e la forma del pericarion, il numero di processi, le loro connessioni sinaptiche, la natura della ramificazione dendritica, le caratteristiche elettrofisiologiche, la chimica dei neurotrasmettitori, la posizione nelle reti funzionali e molte altre caratteristiche. Per questo motivo, le classificazioni dei neuroni sono numerose. Presentiamo alcuni criteri di classificazione ed esempi pertinenti.
Il numero di tiri. Secondo questo criterio, i neuroni si distinguono apolare(nessun processo, neuroblasti precoci), unipolare(l'unico processo, i neuroni pseudo-unipolari dei gangli spinali possono essere considerati formalmente a singolo processo), bipolare(due processi: assone e dendrite, cellule del recettore olfattivo), multipolare(il numero di processi > 2, un assone, il resto sono dendriti, motoneuroni delle corna anteriori del midollo spinale).
Chimica del neurotrasmettitore. Il criterio di classificazione è la sintesi, l'accumulo nelle vescicole sinaptiche e l'escrezione nella fessura sinaptica di uno specifico neurotrasmettitore. Allo stesso tempo, "ergico" viene aggiunto al nome del neurotrasmettitore. A volte viene utilizzato come criterio il tipo di recettore di membrana che rileva la presenza di un neurotrasmettitore (in questo caso si aggiunge “recettivo”). Esempi: colinergico neuroni, adrenergici cellule nervose.
Posizione del neurone v arco riflesso consente di evidenziare sensibile(percependo un segnale dall'ambiente esterno o interno) neuroni, il motore(elementi contrattili e secretori innervanti) neuroni e quelli tra di loro intercalare(associativo nelle reti neurali) cellule nervose (interneuroni).
Direzione dell'eccitazione: al centro - afferente cellule nervose, alla periferia - efferente neuroni.
Modalità- la natura del segnale percepito e trasmesso (ad esempio, meccanorecettore, visivo, olfattivo neuroni, ecc.).
sinapsi
Le sinapsi sono contatti intercellulari specializzati che trasmettono segnali da un neurone all'altro utilizzando neurotrasmettitori. Nella sinapsi si distinguono le parti presinaptiche e postsinaptiche, separate da una fessura sinaptica larga 20-30 nm. I neuroni presinaptici sintetizzano, immagazzinano e secernono neurotrasmettitori. Quando il potenziale di membrana cambia, il neurotrasmettitore viene rilasciato nella fessura sinaptica e si lega ai suoi recettori nella membrana postsinaptica, provocando un cambiamento nel potenziale di membrana del neurone postsinaptico.
Classificazione delle sinapsi. Le sinapsi sono assodendritiche, assosomatiche, asso-assonali e dendrodendritiche.
assodendritico- sinapsi tra l'assone di un neurone ei dendriti di un altro neurone (Fig. 8-15). Una variante sono le sinapsi axo-spiky.
Sinapsi asso-spiky. La maggior parte delle sinapsi eccitatorie sono localizzate in escrescenze di dendriti contenenti un gran numero di actina e chiamate spine.
Asso-assonale sinapsi tra assoni di diversi neuroni.
assosomatico- sinapsi tra i terminali degli assoni di un neurone e il corpo di un altro neurone.
Dendrodendritico sinapsi tra i dendriti dei neuroni. Neurotrasmettitori. La maggior parte dei neurotrasmettitori sono amminoacidi e loro derivati. Alcuni neuroni modificano gli amminoacidi con la formazione di ammine (norepinefrina, serotonina, acetilcolina), altri secernono neurotrasmettitori di natura peptidica (endorfine, encefaline, vari neuropeptidi). Alcuni neuroni usano l'acetilcolina come neurotrasmettitore. Alcuni neuroni possono sintetizzare più di un neurotrasmettitore.
neuroglia
Le cellule neurogliali costituiscono quasi la metà del volume del cervello. Tra le cellule gliali del cervello sono isolate la glia ependimale, la macroglia e la microglia (Fig. 8-16). Le macroglia sono composte da astrociti e oligodendrociti. Nel sistema nervoso periferico ci sono cellule di Schwann e un gruppo di cellule ausiliarie che circondano i neuroni nei gangli.
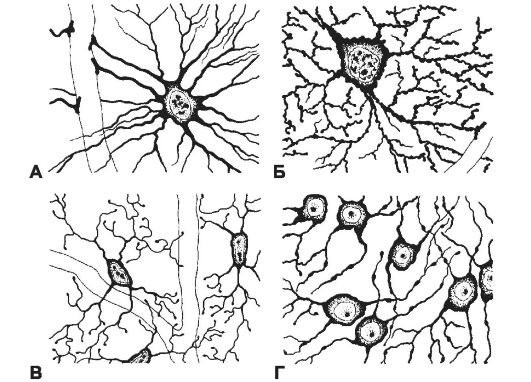
Astrociti. I processi di queste cellule a forma di stella si estendono dal corpo cellulare in diverse direzioni, intrecciano neuroni, vasi sanguigni, cellule (ependimi) dei ventricoli del cervello, formando estensioni sotto forma di una gamba terminale. Marcatore di astrociti - acido fibrillare gliale
 Riso. 8-15. Sinapsi interneuronali.
Riso. 8-15. Sinapsi interneuronali.
lok di filamenti intermedi. Ci sono astrociti fibrosi e protoplasmatici.
Astrociti fibrosi hanno processi lunghi, debolmente o non ramificati; presente nella materia bianca del cervello. Le estensioni terminali dei loro processi circondano le intercettazioni nelle fibre mieliniche.
Astrociti protoplasmatici si trovano in materia grigia cervello. Le cellule hanno numerosi processi brevi e ramificati. Le estensioni finali dei processi circondano le aree delle sinapsi.
Cellule che formano la mielina. La mielina è una struttura compatta di membrane che si avvolgono a spirale attorno agli assoni. Il 70% della massa della mielina è costituito da lipidi. È la mielina che conferisce alla materia bianca il suo caratteristico colore che la distingue dalla materia grigia. Cellule che formano la mielina - Schwann e oligodendrociti.
Oligodendrociti(Fig. 8-16D) tendono ad essere cellule più piccole degli astrociti. Gli oligodendrociti si trovano nella materia grigia del cervello
 Riso. 8-16. Cellule gliali: UN- astrocita fibroso; B- astrocita protoplasmatico; IN- microglia; G- oligodendrogliociti.
Riso. 8-16. Cellule gliali: UN- astrocita fibroso; B- astrocita protoplasmatico; IN- microglia; G- oligodendrogliociti.
in contatto diretto con perikarya e processi di neuroni. Nella materia bianca, gli oligodendrociti si trovano in file tra le fibre nervose. Con l'aiuto di sottili processi non ramificati, gli oligodendrociti contattano gli assoni e, muovendosi rispetto all'assone con le estremità appiattite dei processi, lo circondano con una lamina circolare di mielina (una buona analogia è che ruotando attorno agli assoni, avvolgono la mielina attorno all'assone). Ogni oligodendrocita mielinizza diversi assoni con l'aiuto dei suoi processi.
Cellule di Schwann fanno parte delle fibre nervose periferiche e formano la mielina. La mielinizzazione degli assoni nel sistema nervoso periferico è illustrata nella Figura 8-17. Ogni cellula di Schwann mielinizza un assone. Ependima glia. Le cellule ependimali sono cuboidali e formano uno strato di rivestimento epiteliale canale centrale e ventricoli del cervello. Le cellule hanno ciglia ben sviluppate e formano anche giunzioni intermedie, strette e gap, quindi
formiamo una barriera di permeabilità. Gli ependimociti modificati secernono liquido cerebrospinale. Microglia. Le cellule microgliali (Fig. 8-16B) hanno taglia piccola, forma irregolare, numerosi processi di ramificazione, un nucleo con grossi ciuffi di cromatina, molti lisosomi, granuli di lipofuscina e densi corpi lamellari. La funzione della microglia nel cervello intatto non è chiara.
SISTEMA NERVOSO PERIFERICO
FIBRE NERVOSE
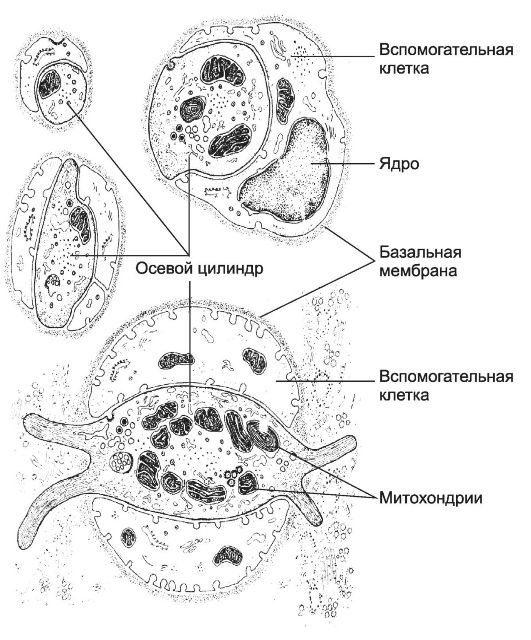
A seconda che le cellule di Schwann formino la mielina attorno al cilindro assiale, vengono isolate le fibre amielinizzate e mielinizzate.
fibre non mielinizzate sono costituiti da assoni di cellule nervose (cilindri assiali) circondati da cellule di Schwann. Quando il cilindro assiale è immerso nella cellula di Schwann, la sua membrana cellulare si chiude e si forma mesassone(Fig. 8-17) - doppie membrane della cellula di Schwann. Ogni gabbia di Schwann circonda allo stesso modo diversi cilindri assiali.
fibra di mielinaè costituito da un cilindro assiale, attorno al quale le cellule di Schwann formano la mielina a causa dell'allungamento e della stratificazione concentrica delle membrane mesassoni. Ogni cellula di Schwann mielinizza un piccolo segmento di un solo assone. La mielina viene interrotta a intervalli regolari intercettazioni nodali. In realtà, questi sono i confini tra due celle di Schwann vicine. nella mielina nervi periferici ci sono piccole illuminazioni - tacche mieliniche. La mielina esterna si trova strato sottile il citoplasma della cellula di Schwann e il suo nucleo.
cilindro dell'assale contiene mitocondri, elementi del reticolo endoplasmatico liscio, vescicole, nonché elementi del citoscheletro: microtubuli, neurofilamenti e microfilamenti. Il diametro dell'assone, e quindi la velocità degli impulsi lungo questo assone, è determinato dal numero di neurofilamenti in esso contenuti.
trasporto di assoni vari componenti forniscono microtubuli di tubulina. Distinguere veloce(100-1000 mm/giorno) e lento trasporto di assoni (1-10 mm/giorno), così come anterogrado(trasporto dal pericarion) e retrogrado(al pericarion). Il materiale principale del trasporto anterogrado sono le proteine sintetizzate nel pericarion (ad esempio proteine del canale ionico, enzimi per la sintesi di neurotrasmettitori).
Intercettazioni nodali. Al confine tra cellule di Schwann vicine, una porzione della membrana plasmatica dell'assone (axolemma) non è ricoperta dalla mielina. Qui le cellule di Schwann formano numerosi processi di intreccio. Ak-
 Riso. 8-17. formazione di fibre di mielina. Nella parte superiore della figura sono mostrati fasi iniziali formazione della mielina. Man mano che il mesassone si allunga, si verifica una stratificazione a spirale della membrana cellulare di Schwann. Allo stesso tempo, il suo citoplasma viene spostato alla periferia. La tacca mielinica è una striscia stretta all'interno della quale le membrane mieliniche divergono e tra di loro ci sono piccole isole del citoplasma delle cellule di Schwann. La parte inferiore della figura mostra un diagramma di una sezione longitudinale di una fibra mielinica nell'area dell'intercettazione nodale - il confine tra cellule di Schwann adiacenti, dove sono collegate intrecciando processi corti. Nell'intercettazione, l'axolemma del cilindro assiale non è ricoperto da una guaina mielinica.
Riso. 8-17. formazione di fibre di mielina. Nella parte superiore della figura sono mostrati fasi iniziali formazione della mielina. Man mano che il mesassone si allunga, si verifica una stratificazione a spirale della membrana cellulare di Schwann. Allo stesso tempo, il suo citoplasma viene spostato alla periferia. La tacca mielinica è una striscia stretta all'interno della quale le membrane mieliniche divergono e tra di loro ci sono piccole isole del citoplasma delle cellule di Schwann. La parte inferiore della figura mostra un diagramma di una sezione longitudinale di una fibra mielinica nell'area dell'intercettazione nodale - il confine tra cellule di Schwann adiacenti, dove sono collegate intrecciando processi corti. Nell'intercettazione, l'axolemma del cilindro assiale non è ricoperto da una guaina mielinica.
l'intercettazione solemma contiene molti canali voltaggio-dipendenti necessari per mantenere l'attività impulsiva.
tacche mieliniche- aree di stratificazione mielinica, formatesi durante la mielinizzazione; contengono il citoplasma delle cellule di Schwann. La funzione delle tacche non è chiara.
nervi periferici sono costituiti da fibre mielinizzate e non mielinizzate raggruppate in fasci e membrane di tessuto connettivo.
Guaine del nervo. Le guaine nervose comprendono l'endoneurio, il perinevrio e l'epinevrio.
♦ Endoneurio- tessuto connettivo lasso tra individuo fibre nervose.
♦ Perinevrio contiene la parte esterna - tessuto connettivo denso che circonda ciascun fascio di fibre nervose e parte interna- diversi strati concentrici di cellule perineurali piatte, ricoperte esternamente ed internamente da una membrana basale eccezionalmente spessa contenente collagene di tipo IV, laminina e fibronectina.
Barriera perineurale necessario per mantenere l'omeostasi nell'endoneurio, forma la parte interna del perinevrio, uno strato di cellule perineurali di tipo epiteliale collegato da giunzioni strette. La barriera controlla il trasporto di molecole attraverso il perinevrio alle fibre nervose e impedisce agli agenti infettivi di entrare nell'endoneurio.
♦ epinevrio- tessuto connettivo fibroso che unisce tutti i fasci nella composizione del nervo.
Riserva di sangue. Il nervo periferico contiene una vasta rete di vasi sanguigni. Nell'epineurio e nella parte esterna (tessuto connettivo) del perinevrio - arteriole e venule, nonché vasi linfatici. L'endoneurio contiene capillari sanguigni.
Innervazione. Il nervo periferico ha fibre nervose speciali - nervi nervosi- sottili fibre nervose sensoriali e simpatiche. La loro fonte: il nervo stesso o il plesso vascolare.
Degenerazione e rigenerazione dei nervi. Quando un nervo è danneggiato, il segmento centrale (associato al perikarya) e il segmento periferico (distale rispetto al sito della lesione) subiscono modifiche diverse. La degenerazione delle fibre nervose (Fig. 8-18) si verifica su una piccola estensione del segmento centrale e su tutto il segmento periferico. Successivamente, si verifica la rigenerazione del segmento periferico. Le cellule di Schwann svolgono un ruolo di primo piano in questo processo. Le cellule di Schwann stimolano l'allungamento dell'assone e controllano la sua crescita diretta verso il bersaglio. In assenza di cellule di Schwann, gli assoni non possono crescere su distanze significative.
NODI NERVI SENSIBILI
nodo spinale (ganglio spinale) ricoperto da una capsula di tessuto connettivo. All'interno del nodo ci sono gruppi di neuroni sensoriali pseudo-unipolari, tra i quali ci sono fasci di mielina

Riso. 8-18. Rigenerazione nervosa. UN- fibra prima del danno; B- nel segmento periferico, l'assone degenera, le cellule proliferano nel sito del danno; IN- rigenerazione degli assoni nel segmento centrale, germinazione dei rami degli assoni nel segmento periferico; G- rigenerazione completa della fibra nervosa e ripristino delle connessioni.
fibre. I pericarioni dei neuroni sono arrotondati e circondati da cellule satellite. La popolazione dei neuroni gangliari è eterogenea. I nodi spinali contengono oltre 20 diversi sottotipi di neuroni sensoriali.
TERMINAZIONI NERVOSE
Le terminazioni degli assoni dei nervi periferici sono divise in sensibile(afferente) e il motore(efferente).
Terminazioni nervose sensoriali
Gli stimoli esterni, così come i segnali sullo stato dell'ambiente interno e dei muscoli del corpo, registrano terminazioni nervose sensibili - recettori sensoriali. Distinguere tra liberi e non liberi
dispositivi percettivi, e tra questi ultimi corpi incapsulati.
Gratuito le terminazioni nervose sono rami terminali del processo periferico di un neurone sensibile.
Non gratis le terminazioni nervose, oltre ai rami terminali, contengono cellule speciali. Quasi tutte le terminazioni di questo tipo sono meccanocettori.
Incapsulato i meccanorecettori (corpi incapsulati) sono terminazioni non libere che hanno una capsula di tessuto connettivo formata.
terminazioni nervose libere(il fico. 8-21) si calmano in strati intermedi tessuto connettivo organi interni e pelle, così come negli strati basale e spinoso dell'epidermide, e nelle aree della pelle con elevata sensibilità tattile, raggiungono lo strato granulare. La maggior parte delle terminazioni nervose libere sono meccanocettori. Alcune terminazioni nell'epidermide sono chiaramente specializzate per registrare i cambiamenti di temperatura. Esistono anche recettori in grado di rilevare variazioni di pH, pO 2 e pCO 2.
Complesso di epiteliociti tattili con terminazione nervosa. Gli epiteliociti tattili sono cellule arrotondate o allungate situate principalmente nell'epitelio della pelle delle dita, delle labbra e dei genitali esterni (Fig. 8-22). Queste cellule formano contatti con le terminazioni nervose. Nell'area di contatto, il terminale è nettamente espanso, contiene molti mitocondri, neurofilamenti e microtubuli. Le cellule epiteliali tattili sono più grandi dei cheratinociti vicini; il citoplasma contiene quantità moderate di mitocondri, lisosomi, corpi multivescicolari, vacuoli e microfilamenti. Le cellule epiteliali tattili sono caratterizzate da specifici granuli osmiofili dimensioni da 80 a 200 nm. Sono concentrati principalmente nelle parti del citoplasma rivolte verso la terminazione nervosa. Peptidi e sostanze specifiche dei neuroni (ad esempio metionina-encefalina, VIP, sostanza P) sono stati trovati negli epiteliociti tattili, il che indica la funzione endocrina delle cellule e ci consente di considerarli come un componente del sistema neuroendocrino diffuso. Gli epiteliociti tattili sono coinvolti nel riconoscimento della forma di un oggetto, dei suoi bordi e della struttura della superficie.
corpi lamellari(Fig. 8-24) trovato nel tessuto connettivo della pelle e vari corpi, hanno una forma ovale, dimensioni fino a 0,5-1,0 mm. Il bulbo interno, la capsula esterna e la fibra nervosa terminale sono i componenti principali del corpuscolo.
Un'adeguata stimolazione meccanica dei corpi lamellari porta alla comparsa di un potenziale recettore nella parte terminale della terminazione nervosa. Raggiunto un valore critico, il potenziale del recettore nella prima intercettazione nodale provoca la comparsa di potenziali d'azione. La capsula esterna del recettore è un filtro che lascia passare solo la componente dinamica dell'impatto meccanico.
 Riso. 8-21. Terminazioni nervose libere nel tessuto connettivo. Il cilindro assiale è a contatto con le celle accessorie per una notevole distanza o ne è completamente circondato. La terminazione nervosa libera mostrata nella parte inferiore della figura ha un'organizzazione bilaterale: al centro si trova una terminazione nervosa ellittica ricoperta di cellule accessorie.
Riso. 8-21. Terminazioni nervose libere nel tessuto connettivo. Il cilindro assiale è a contatto con le celle accessorie per una notevole distanza o ne è completamente circondato. La terminazione nervosa libera mostrata nella parte inferiore della figura ha un'organizzazione bilaterale: al centro si trova una terminazione nervosa ellittica ricoperta di cellule accessorie.
Corpi tattili(Fig. 8-26) sono presenti nello strato papillare della pelle, localizzato in modo più compatto sulla punta delle dita. Il corpo è allungato. Il nucleo del corpuscolo è formato da cellule accessorie lamellari, alternate a terminazioni nervose espanse.
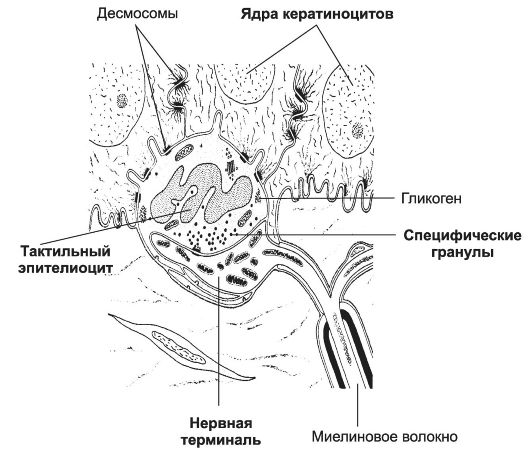 Riso. 8-22. Complesso di epiteliociti tattili con terminazione nervosa. L'epiteliocita tattile si trova nello strato basale dell'epidermide, forma processi e si lega ai cheratinociti usando i desmosomi. Il terminale nervoso espanso entra in contatto con la cellula epiteliale tattile. Granuli specifici si trovano nella parte della cellula rivolta verso la terminazione nervosa.
Riso. 8-22. Complesso di epiteliociti tattili con terminazione nervosa. L'epiteliocita tattile si trova nello strato basale dell'epidermide, forma processi e si lega ai cheratinociti usando i desmosomi. Il terminale nervoso espanso entra in contatto con la cellula epiteliale tattile. Granuli specifici si trovano nella parte della cellula rivolta verso la terminazione nervosa.
Altri corpi(p. es., corpi fusiformi e genitali, fiaschi di Krause) sono meno comuni.
Recettori muscolari e articolari. Questi includono fusi muscolari, organi tendinei e terminazioni nervose sensoriali nella capsula articolare.
fusi muscolari(fig. 8-27, vedi anche fig. 7-9) - dispositivi sensibili di rilevamento muscolo scheletrico. I principali elementi strutturali del fuso muscolare sono le fibre muscolari intrafusali, le fibre nervose e una capsula. Il fuso muscolare contiene da 1 a 10 fibre muscolari intrafusali corte. Nella loro parte equatoriale, i nuclei formano un ammasso compatto (fibre con
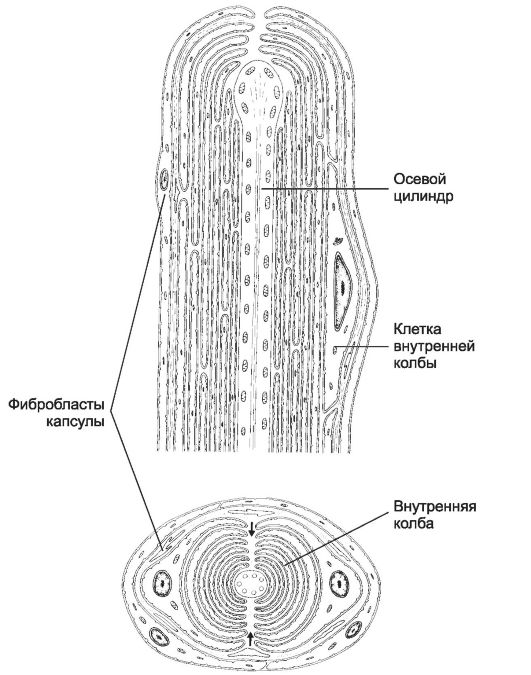 Riso. 8-24. corpo lamellareè costituito da una fiasca interna organizzata bilateralmente e da una capsula esterna di tessuto connettivo. Le cellule della fiasca interna formano processi che circondano il terminale nervoso sensibile in semianelli concentrici. Le frecce indicano la fenditura della fiasca interna, che corre parallela all'asse corto dell'ellisse della terminazione nervosa.
Riso. 8-24. corpo lamellareè costituito da una fiasca interna organizzata bilateralmente e da una capsula esterna di tessuto connettivo. Le cellule della fiasca interna formano processi che circondano il terminale nervoso sensibile in semianelli concentrici. Le frecce indicano la fenditura della fiasca interna, che corre parallela all'asse corto dell'ellisse della terminazione nervosa.
 Riso. 8-26. Corpo tattile circondato da una capsula di tessuto connettivo con fibre di collagene del tessuto connettivo circostante intrecciate in esso. Le fibre nervose che si avvicinano al corpo perdono la mielina e vanno all'interno del corpo, dove formano numerosi terminali circondati da cellule ausiliarie.
Riso. 8-26. Corpo tattile circondato da una capsula di tessuto connettivo con fibre di collagene del tessuto connettivo circostante intrecciate in esso. Le fibre nervose che si avvicinano al corpo perdono la mielina e vanno all'interno del corpo, dove formano numerosi terminali circondati da cellule ausiliarie.
borsa nucleare) o disposti in una catena (fibre con una catena nucleare). I terminali delle fibre Ια formano una spirale all'interno della zona equatoriale di entrambi i tipi di fibre muscolari intrafusali (terminazioni primarie o anulospirali). I terminali delle fibre II più sottili terminano sulle fibre intrafusali nella zona adiacente a quella equatoriale.
organi tendinei situato nella parte terminale del tendine al confine con il muscolo, nonché nei legamenti della capsula articolare. Il recettore ha una forma a fuso ed è circondato da una capsula costituita da diversi strati. cellule piatte. La capsula del recettore è una continuazione del perinevrio e contiene capillari. I terminali delle fibre mieliniche afferenti (Ib) partecipano alla formazione dell'organo tendineo; si ramificano tra fasci di fibre collagene a spirale situate nello spazio pieno di liquido.
Terminazioni nervose sensoriali della capsula articolare- elemento importante sistema propriocettivo del corpo. Corpi fusiformi
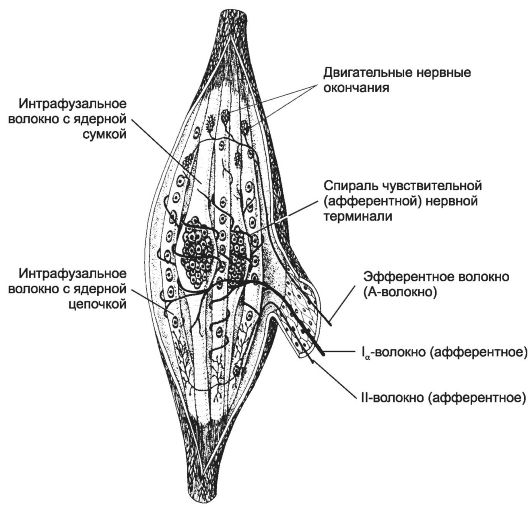 Riso. 8-27. Fuso muscolare. Le fibre muscolari intrafusali con un accumulo compatto di nuclei sono fibre con un sacco nucleare; nelle fibre intrafusali con una catena nucleare, i nuclei sono distribuiti più uniformemente lungo la lunghezza della fibra. Le terminazioni primarie (annulospirali) si trovano nella regione equatoriale. Più vicino alle estremità delle fibre intrafusali ci sono i terminali delle sottili fibre afferenti II - le terminazioni secondarie. Le fibre efferenti formano sinapsi neuromuscolari con fibre intrafusali alla loro estremità.
Riso. 8-27. Fuso muscolare. Le fibre muscolari intrafusali con un accumulo compatto di nuclei sono fibre con un sacco nucleare; nelle fibre intrafusali con una catena nucleare, i nuclei sono distribuiti più uniformemente lungo la lunghezza della fibra. Le terminazioni primarie (annulospirali) si trovano nella regione equatoriale. Più vicino alle estremità delle fibre intrafusali ci sono i terminali delle sottili fibre afferenti II - le terminazioni secondarie. Le fibre efferenti formano sinapsi neuromuscolari con fibre intrafusali alla loro estremità.
situato nelle regioni periferiche della capsula. Piccoli corpi lamellari, nonché terminazioni nervose libere - terminali di sottili fibre mielinizzate e fibre non mielinizzate, tra le quali, a quanto pare, sono presenti anche i recettori del dolore, sono ampiamente rappresentati in tutte le componenti dell'articolazione, ma raggiungono la massima densità nel menisco e disco articolare.
terminazioni nervose motorie
Le caratteristiche delle terminazioni nervose motorie sono discusse di seguito sull'esempio delle sinapsi neuromuscolari. Come in altre sinapsi, ci sono regioni presinaptiche e postsinaptiche separate da una fessura sinaptica.
zona presinaptica. La terminazione nervosa motoria è ricoperta esternamente da una cellula di Schwann, ha un diametro di 1-1,5 micron e forma la regione presinaptica della sinapsi neuromuscolare (Fig. 8-29).
 Riso. 8-29. sinapsi neuromuscolare. La parte presinaptica è formata dal terminale assone del motoneurone e contiene un accumulo di vescicole sinaptiche vicino alla membrana presinaptica, così come i mitocondri. Le pieghe postsinaptiche aumentano la superficie della membrana postsinaptica. Nella fessura sinaptica c'è una membrana basale sinaptica (una continuazione della membrana basale della fibra muscolare), entra nelle pieghe postsinaptiche. La fessura sinaptica contiene anche molecole di acetilcolinesterasi. Questo enzima scompone l'acetilcolina ed elimina l'effetto del segnale depolarizzante sulla fibra muscolare.
Riso. 8-29. sinapsi neuromuscolare. La parte presinaptica è formata dal terminale assone del motoneurone e contiene un accumulo di vescicole sinaptiche vicino alla membrana presinaptica, così come i mitocondri. Le pieghe postsinaptiche aumentano la superficie della membrana postsinaptica. Nella fessura sinaptica c'è una membrana basale sinaptica (una continuazione della membrana basale della fibra muscolare), entra nelle pieghe postsinaptiche. La fessura sinaptica contiene anche molecole di acetilcolinesterasi. Questo enzima scompone l'acetilcolina ed elimina l'effetto del segnale depolarizzante sulla fibra muscolare.
La regione presinaptica è ricca di vescicole sinaptiche e mitocondri.
membrana presinaptica- parte specializzata dell'axolemma della terminazione nervosa. Nella membrana presinaptica sono state identificate le cosiddette zone attive, aree di ispessimento della membrana che sono direttamente correlate alla secrezione del neurotrasmettitore. Vicino alle zone attive nella membrana presinaptica ci sono depressioni, il cui numero è correlato al livello di secrezione di acetilcolina. La membrana presinaptica contiene canali Ca 2 + dipendenti dalla tensione. Quando la membrana è depolarizzata, i canali si aprono e gli ioni Ca 2 + entrano nel terminale, innescando la secrezione di acetilcolina.
vescicole sinaptiche sono presenti in quasi tutte le aree della terminazione nervosa, ma nelle immediate vicinanze della membrana presinaptica formano un accumulo pronunciato. Le dimensioni delle bolle variano, il loro diametro medio è di 50 nm. Nelle sinapsi colinergiche, le vescicole sinaptiche leggere contengono acetilcolina.
regione postsinaptica. La regione postsinaptica è caratterizzata dalla presenza di grandi mitocondri con creste ben sviluppate e un gran numero di ribosomi.
membrana postsinaptica- una parte specializzata del plasmolemma delle fibre muscolari - forma numerose intussuscezioni, dalle quali le pieghe postsinaptiche si estendono fino a una profondità di 0,5-1,0 micron, il che aumenta significativamente l'area della membrana. I recettori N-colinergici sono incorporati nella membrana postsinaptica, la loro concentrazione raggiunge 20-30 mila per 1 micron 2.
fessura sinaptica. La membrana basale sinaptica passa attraverso la fessura sinaptica, trattiene il terminale dell'assone nell'area della sinapsi e controlla la posizione dei recettori colinergici nella membrana postsinaptica.
SISTEMA AUTONOMICO
Neuroni reparto parasimpatico originano dalla cresta neurale a livello di 1-7 somiti (dipartimento nervo vago) e caudale al somite 28 (lombosacrale). I neuroni della divisione simpatica e le cellule cromaffini del midollo surrenale si sviluppano dalla cresta neurale a livello dei somiti 8-28. L'innervazione vegetativa (simpatica e parasimpatica) di ciascun organo interno è discussa nei capitoli corrispondenti del manuale. Qui presentiamo uno schema generale delle connessioni anatomiche della divisione simpatica (Fig. 8-32).
La struttura delle parti periferiche del sistema nervoso autonomo sarà considerata utilizzando l'esempio dell'innervazione autonomica tratto digerente(Vedere anche la sezione "Apparato nervoso dell'apparato digerente" nel capitolo 12, figure 12-19 e figure 19-20). Nel muro tubo digerente ci sono plessi nervosi contenenti gangli. In quest'ultimo si concentrano i perikarya dei neuroni autonomi. Il numero di neuroni in un ganglio varia da pochi a centinaia. L'insieme degli elementi nervosi del tubo digerente costituisce il sistema nervoso enterico.
 Riso. 8-32. Reparto simpatico sistema nervoso autonomo. I pericarion del primo neurone si trovano nelle colonne laterali del midollo spinale. I loro assoni passano come parte delle radici anteriori e del ramo di collegamento bianco e terminano sul perikarya del secondo neurone nei gangli paravertebrali della catena simpatica, nei gangli prevertebrali e nei gangli (terminali) situati vicino agli organi da essi innervati. Gli assoni del secondo neurone dei gangli paravertebrali, prevertebrali e terminali terminano in organi interni, pelle, parete dei vasi sanguigni. Gli assoni di alcuni neuroni dei gangli paravertebrali passano attraverso il ramo di collegamento grigio.
Riso. 8-32. Reparto simpatico sistema nervoso autonomo. I pericarion del primo neurone si trovano nelle colonne laterali del midollo spinale. I loro assoni passano come parte delle radici anteriori e del ramo di collegamento bianco e terminano sul perikarya del secondo neurone nei gangli paravertebrali della catena simpatica, nei gangli prevertebrali e nei gangli (terminali) situati vicino agli organi da essi innervati. Gli assoni del secondo neurone dei gangli paravertebrali, prevertebrali e terminali terminano in organi interni, pelle, parete dei vasi sanguigni. Gli assoni di alcuni neuroni dei gangli paravertebrali passano attraverso il ramo di collegamento grigio.
neuroni autonomi. Ci sono tre tipi di neuroni nel sistema nervoso enterico.
Neuroni enterici di tipo I: perikarya hanno una forma appiattita, un lungo assone e un gran numero di corti dendriti con una base espansa.
Neuroni enterali tipo II avere un pericarion di forma ovale con una superficie liscia e lunghi tiri.
Neuroni enterici di tipo III: perikarya hanno una forma ovale o irregolare, un lungo assone e un gran numero di dendriti relativamente corti di varie lunghezze.
Connessioni di neuroni. I neuroni di tipo I formano sinapsi con le SMC e con i neuroni di tipo II. Si ritiene che i neuroni di tipo I siano motoneuroni e che i neuroni di tipo II siano sensibili. I processi dei neuroni di tipo III non solo entrano in contatto con i neuroni dei gangli vicini, ma penetrano anche nelle mucose e sottomucose.
sinapsi. L'assone del neurone autonomo postgangliare forma numerosi ispessimenti locali - vene varicose contenenti vescicole sinaptiche (vedi Figura 7-29). Questi ispessimenti sono siti di secrezione di neurotrasmettitori.
I neuroni autonomi sintetizzano e secernono vari biologicamente sostanze attive(neurotrasmettitori e modulatori). neurotrasmettitori(acetilcolina, norepinefrina e serotonina) provocano contrazione/rilassamento del SMC, eccitazione/inibizione dei neuroni del sistema nervoso enterico, potenziamento/soppressione della secrezione delle ghiandole esocrine e delle cellule enteroendocrine. In alcuni neuroni dei gangli del plesso intermuscolare, la serotonina e la sostanza P sono presenti insieme.
SISTEMA NERVOSO CENTRALE
La sezione della testa del tubo neurale forma tre estensioni, o vescicole cerebrali primarie: proencefalo (prosencefalo)mesencefalo(mesencefalo) E cervello romboidale(rombencefalo). Alla settimana 5, il cervello anteriore è diviso in due parti: telencefalo(telencefalo), formato dagli emisferi primari del cervello, e diencefalo(diencefalo) con i bulbi oculari. Media cervello e a forma di diamante il cervello è separato da un istmo profondo. Il cervello romboidale è diviso in rombencefalo(metencefalo) E midollo(mielencefalo). Sviluppa dal rombencefalo ponte(ponte cerebrale) E cervelletto(cervelletto). Il resto delle forme del tubo neurale midollo spinale.
Midollo spinale
Il midollo spinale è costituito da due metà simmetriche (Fig. 8-35), collegate da uno stretto ponte contenente il canale centrale, residuo della cavità del tubo neurale. SU sezione trasversale facilmente distinguibile sdraiato all'esterno materia bianca e una massa oscura più profonda - Materia grigia.
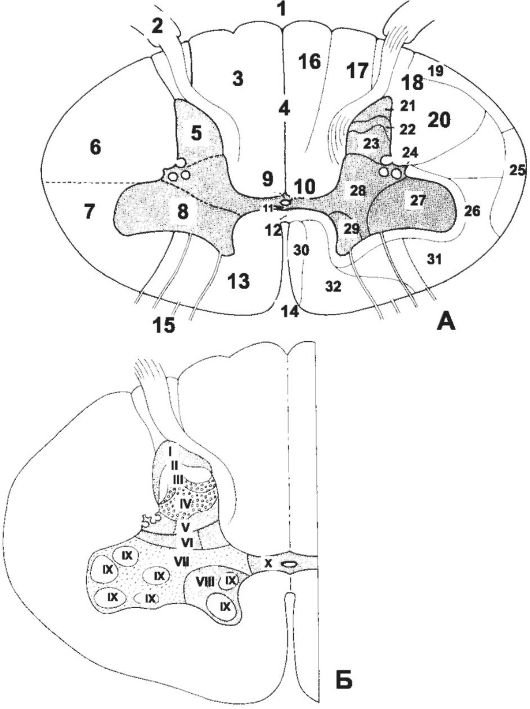 Riso. 8-35. Sezione trasversale del midollo spinale. UN- strutture di materia grigia e bianca. B- topografia delle placche in materia grigia. 1 - Solco mediano posteriore; 2 - Radice dorsale; 3 - Funicolo posteriore; 4 - Setto mediano posterio; 5 - corno posterio; 6 - Funicolo posterolaterale; 7 - Funicolo anterolaterale;
8 - corno anteriore; 9 - Commissura grisea posteriore; 10 - Canalis centralis; 11 - Commissura grisea anteriore; 12 - Commissura alba; 13 - Funicolo anteriore; 14 - Fissura mediana anteriore; 15 - radice ventrale; 16 - Fascicolo gracile; 17 - Fascicolo cuneo; 18 - Fascicolo dorsolaterale; 19 - Tractus spinocerebellaris posteriore; 20 - tratto piramidale laterale; 21 - cellule marginali; 22 - sostanza gelatinosa; 23 - N. proprio; 24 - Processo reticolare; 25 - Tractus spinocerebellaris anteriore; 26 - Fascicoli propri; 27 - Cellulae motoriae laterale; 28 - Sostanza intermedia; 29 - Cellule motorie mediali; 30 - Tractus piramidale anteriore; 31 - Fasciculus anterolateralis: tractus spinotectalis, tractus spinothalamicus, tractus spinoannularis, tractus spino-olivaris; 32 - Fascicolo longitudinale mediale: tractus vestibulospinalis medialis, tractus vestibulospinalis lateralis, tractus reticulospinalis, tractus tectospinalis, tractus interstitiospinalis.
Riso. 8-35. Sezione trasversale del midollo spinale. UN- strutture di materia grigia e bianca. B- topografia delle placche in materia grigia. 1 - Solco mediano posteriore; 2 - Radice dorsale; 3 - Funicolo posteriore; 4 - Setto mediano posterio; 5 - corno posterio; 6 - Funicolo posterolaterale; 7 - Funicolo anterolaterale;
8 - corno anteriore; 9 - Commissura grisea posteriore; 10 - Canalis centralis; 11 - Commissura grisea anteriore; 12 - Commissura alba; 13 - Funicolo anteriore; 14 - Fissura mediana anteriore; 15 - radice ventrale; 16 - Fascicolo gracile; 17 - Fascicolo cuneo; 18 - Fascicolo dorsolaterale; 19 - Tractus spinocerebellaris posteriore; 20 - tratto piramidale laterale; 21 - cellule marginali; 22 - sostanza gelatinosa; 23 - N. proprio; 24 - Processo reticolare; 25 - Tractus spinocerebellaris anteriore; 26 - Fascicoli propri; 27 - Cellulae motoriae laterale; 28 - Sostanza intermedia; 29 - Cellule motorie mediali; 30 - Tractus piramidale anteriore; 31 - Fasciculus anterolateralis: tractus spinotectalis, tractus spinothalamicus, tractus spinoannularis, tractus spino-olivaris; 32 - Fascicolo longitudinale mediale: tractus vestibulospinalis medialis, tractus vestibulospinalis lateralis, tractus reticulospinalis, tractus tectospinalis, tractus interstitiospinalis.
MATERIA GRIGIA
La materia grigia è costituita da processi di cellule nervose e il loro perikarya, formando grappoli - nucleo, uniti dentro record. Ogni metà della materia grigia forma sporgenze in tutto il midollo spinale - grigio poli: posta anteriore - colonna anteriore, palo posteriore - colonna posteriore e palo laterale colonna laterale. La colonna nella sezione trasversale è chiamata corno, rispettivamente, la parte anteriore (corno anteriore), posteriore (cornu posterio) e laterale (cornu laterale). I pericarioni dei neuroni della materia grigia lungo la lunghezza del midollo spinale sono mappati su dieci piastre (vedi Fig. 8-35). La topografia dei nuclei corrisponde alla topografia delle placche, anche se non sempre coincidono.
Neuroni
Nella materia grigia del midollo spinale si trovano i corpi dei neuroni motori, intercalari e autonomi. Ci sono motoneuroni α (grandi) e motoneuroni γ (piccoli).
α -Motoneuroni fanno parte dei nuclei mediale e laterale. Questo è il massimo grandi cellule midollo spinale. I loro assoni formano sinapsi neuromuscolari (Fig. 8-29) con fibre muscolari scheletriche striate.
γ -Motoneuroni innervano le fibre intrafusali dei fusi muscolari (Fig. 8-27, vedi anche Fig. 7-9).
neuroni autonomi situato nei nuclei viscerali delle regioni toracica e lombare (n. intermediomediale), e anche dentro regione sacrale(nuclei parasimpatici sacrali) della zona intermedia (sostanza intermedia) materia grigia. Neuroni N. intermediomediale formano contatti sinaptici con fibre afferenti di sensibilità viscerale generale (afferenze viscerali generali) e i loro assoni entrano in contatto con i neuroni N. intermediolaterale.
MATERIA BIANCA
La materia bianca è costituita da fibre nervose e cellule neurogliali. Le corna della sostanza grigia dividono la sostanza bianca in tre funicoli (posteriore, laterale e anteriore). Condurre percorsi sono formati da una catena di neuroni collegati in serie dai loro processi e forniscono la conduzione dell'eccitazione da neurone a neurone (da nucleo a nucleo). Ci sono percorsi ascendenti, discendenti e misti.
percorsi ascendenti
Le vie ascendenti sono formate dai processi centrali dei neuroni sensoriali dei nodi spinali e dagli assoni dei neuroni intercalari. Processi centrali dei neuroni sensoriali i nodi spinali formano percorsi di sensibilità propriocettiva e tattile, vale a dire un fascio sottile (fascicolo gracile) e fascio a forma di cuneo (fascicolo cuneo).
Assoni di interneuroni proiezione della forma vie afferenti spinotalamico, spinocerebellare, spinolivare, dorso-opercolare, spinoreticolare.
Tratto spinotalamico ventrale(tractus spinothalamicus ventralis) passa nel funicolo anteriore e conduce gli impulsi dai meccanocettori della pelle.
Tratto spinotalamico laterale(tratto spinotalamico laterale) passa nel funicolo laterale, questo è il percorso principale del dolore e della sensibilità alla temperatura. Trasmissione degli impulsi del dolore. La sostanza P è coinvolta nella trasmissione degli stimoli dolorosi come neurotrasmettitore eccitatorio nelle sinapsi tra i processi centrali dei neuroni sensoriali del ganglio spinale e il perikarya dei neuroni del tratto spinotalamico. Il blocco della secrezione della sostanza P e l'alleviamento del dolore sono realizzati attraverso i recettori del peptide oppioide incorporati nella membrana del terminale del processo centrale del neurone sensibile. La fonte del peptide oppioide encefalina è un neurone intercalare.
Tratto spinale anteriore(tratto spinocerebellare anteriore) E tratto dorsale posteriore(tratto spinocerebellare posteriore). Le informazioni su tutti i segnali afferenti di sensibilità profonda e su tutti i cambiamenti nel tono muscolare entrano nel cervelletto attraverso le vie spinocerebellari.
percorso dorsale(tratto spinotectale)- proiezione percorso ascendente sensibilità generale.
Via spinoreticolare(tratto spinoreticolare)- strada di ricevuta di informazioni afferent di riflessi somatici e viscerali.
percorsi discendenti
Le vie discendenti (sopraspinali e spinali) terminano esclusivamente sui motoneuroni.
Tratti sovraspinali condurre segnali da strutture situate sopra il midollo spinale. Questi includono percorsi piramidali(tratto corticospinale anteriore (tratto corticospinalis ventralis; tratto corticospinale laterale, tractus corticospinalis lateralis).via tettospinale(cover-spinale, tractus tectospinalis),tratto nucleare-spinale rosso(tratto rubrospinale, proiezione verso il basso percorso del sistema extrapiramidale),tratto reticolospinale(tratto reticolospinale, efferente percorso del sistema extrapiramidale, controlla il tono muscolare scheletrico e viscerale funzioni motorie), tratto vestibolocerebrale(tratto vestibolospinale),tratto olivo-spinale(tratto olivospinale).tratti spinali situato all'interno del midollo spinale.
Nelle corde posteriori sono presenti fibre discendenti dei processi centrali dei neuroni sensoriali dei nodi spinali: nei segmenti cervicale e toracico superiore si formano fascio a forma di virgola(fascicolo interfascicolare), e dentro lombare - fascio settomarginale(fasciculus septomarginalis).
Nelle corde anteriori del cervicale e toracico fibre del midollo spinale della forma del tratto vestibolare mediale solco-
fascio dchato-marginale(fasciculus sulcomarginalis), controllando il tono dei muscoli del collo secondo varie disposizioni teste.
percorsi misti
A bloccanti misti e percorsi discendenti includono la cintura marginale, i fasci intrinseci del midollo spinale e il generatore del modello centrale.
Generatore di pattern centrale- la rete locale neuroni all'interno del midollo spinale, che controllano e coordinano un ciclo complesso di movimenti stereotipati quando si cammina, si corre, si salta, si nuota, ecc.
Cervello
CERVELLETTO
Cervelletto (cervelletto) coordina i movimenti e l'equilibrio, si trova sopra il midollo allungato e il ponte ed è collegato al tronco encefalico da tre paia di zampe, lungo le quali passano le vie afferenti ed efferenti. Nelle profondità della sostanza bianca del cervelletto giacciono gruppi di neuroni: i nuclei del cervelletto. Le circonvoluzioni del cervelletto, che formano una figura di un albero ramificato su un taglio (arbor vitae), separati da profondi solchi.
Corteccia cerebellare
Ogni giro contiene una stretta lamina di sostanza bianca, completamente ricoperta di materia grigia (corteccia cerebellare, Fig. 8-37), in cui si distinguono tre strati: esterno - molecolare, medio - gangliare e interno - granulare.
strato gangliare, o uno strato di neuroni a forma di pera (strato di cellule di Purkinje) forma i corpi di grandi cellule nervose. I loro perikarya si trovano approssimativamente allo stesso livello dalla superficie della corteccia. 2-3 dendriti fortemente ramificati si estendono dal corpo allo strato molecolare. Attraverso lo strato granulare nella sostanza bianca, un assone parte dal corpo dei neuroni a forma di pera. Gli assoni dei neuroni a forma di pera sono le uniche fibre efferenti che emergono dalla corteccia cerebellare. Terminano sui neuroni dei nuclei cerebellari. Vicino al corpo cellulare, i collaterali partono dall'assone, tornando allo strato gangliare e alle parti profonde dello strato molecolare. Sui neuroni a forma di pera, in un modo o nell'altro, terminano tutti i percorsi afferenti del cervelletto.(Figura 8-37). strato molecolare contiene cesto e cellule stellate. celle a canestro formano numerosi dendriti lunghi e relativamente poco ramificati. Il loro assone è diretto parallelamente alla superficie del cervelletto sullo stesso piano della ramificazione dei dendriti dei neuroni a forma di pera. In tutto l'assone si formano rami, che terminano sotto forma di cestini sui corpi dei neuroni a forma di pera. L'attività dei neuroni a canestro provoca l'inibizione dei neuroni piriformi.
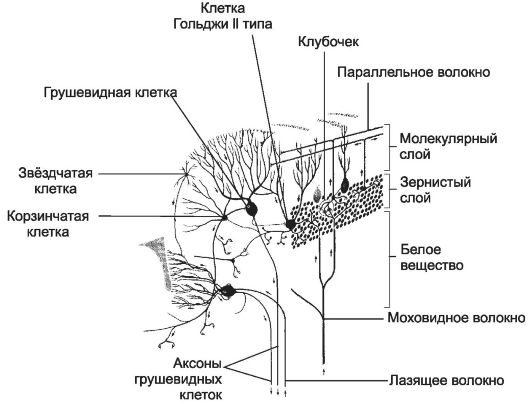 Riso. 8-37. Organizzazione della corteccia cerebellare. La corteccia è formata da tre strati: esterno - molecolare, medio - gangliare e interno - granulare. Dalla sostanza bianca sottostante, le fibre afferenti rampicanti e briofite passano nella corteccia e gli assoni delle cellule a forma di pera escono. Le frecce indicano la direzione di propagazione dell'eccitazione.
Riso. 8-37. Organizzazione della corteccia cerebellare. La corteccia è formata da tre strati: esterno - molecolare, medio - gangliare e interno - granulare. Dalla sostanza bianca sottostante, le fibre afferenti rampicanti e briofite passano nella corteccia e gli assoni delle cellule a forma di pera escono. Le frecce indicano la direzione di propagazione dell'eccitazione.
cellule stellate situato più vicino alla superficie della corteccia. I loro assoni formano contatti sinaptici con i dendriti dei neuroni a forma di pera. Questi neuroni, insieme alle cellule canestro, formano un sistema di neuroni intercalari che trasmettono impulsi inibitori ai dendriti e ai corpi delle cellule a forma di pera in un piano trasversale al giro.
Strato granulare contiene cellule granulari, cellule stellate del Golgi e neuroni orizzontali fusiformi.
Cellule di grano. Il loro corpo è molto piccolo e quasi completamente occupato dal nucleo. 3-4 dendriti molto corti formano rami terminali simili a zampe di uccello. Gli assoni delle cellule granulari salgono nello strato molecolare, dove formano rami a forma di T che corrono paralleli alla superficie del cervelletto in un piano coincidente con la direzione del giro. Questo fibre parallele, formando sinapsi con i dendriti: neuroni a forma di pera, cellule a canestro, cellule stellate, cellule del Golgi di tipo II.
cellule stellate del Golgi. Ci sono neuroni stellati con assoni corti e lunghi. Grandi pericarioni di neuroni stellati con assoni corti spesso si trovano direttamente sotto lo strato gangliare e la maggior parte dei dendriti si diramano nello strato molecolare e formano sinapsi con fibre parallele - assoni di cellule granulari. I loro assoni corti, essendo parte dei glomeruli del cervelletto (glomeruli cerebellosi), terminano con contatti sinaptici su rosette di fibre muschiose.
Neuroni orizzontali fusiformi hanno un piccolo pericarion allungato, da cui si estendono lunghi dendriti orizzontali, che terminano con uno strato di neuroni a forma di pera e uno strato granulare. Gli assoni di questi neuroni formano collaterali nello strato granulare e vanno nella sostanza bianca.
Afferenze cerebellari
La corteccia cerebellare contiene numerose fibre da vari reparti cervello. Nello strato granulare ci sono fibre muschiose. Le fibre rampicanti terminano nello strato molecolare sui dendriti dei neuroni a forma di pera.
fibre di muschio, penetrati nello strato granulare, si ramificano formando rosette terminali che entrano in contatto con i dendriti delle cellule granulari dei glomeruli cerebellari. Le fibre muschiose formano anche sinapsi con i dendriti delle cellule del Golgi di tipo II. Di conseguenza, le fibre muschiose entrano in contatto sia con gli assoni corti dei neuroni stellati dello strato granulare che con i loro dendriti.
fibre rampicanti si avvicinano ai corpi dei neuroni a forma di pera e qui si dividono in diversi rami sottili che intrecciano i dendriti. C'è una fibra rampicante per neurone a forma di pera.
CORTECCIA DEL CERVELLO
Abbaio grande cervello (neocorteccia) contiene 6 strati di cellule nervose
Strati della corteccia
Dall'esterno verso l'interno, si trovano gli strati della corteccia prossimo ordine: molecolare, granulare esterno, piramidale esterno, granulare interno, piramidale interno (gangliare) e polimorfo (multiforme).
IO. Molecolare. Contiene rari perikarion, assoni e dendriti che passano qui.
II. Granulare esterno. Lo strato granulare esterno contiene piccoli neuroni piramidali e stellati.
III. Piramidale esterno. Rappresentato da numerosi neuroni piramidali di medie dimensioni; la dimensione del loro pericarion aumenta nelle parti profonde dello strato.
IV. Granulare interno. Contiene piccole cellule stellate.
v. Piramidale interna(gangliare). Consiste di grandi neuroni piramidali e un piccolo numero di cellule stellate.
VI. Polimorfico.È formato da molti neuroni di varie dimensioni e forme, nonché da un certo numero di neuroni piramidali e granulari.
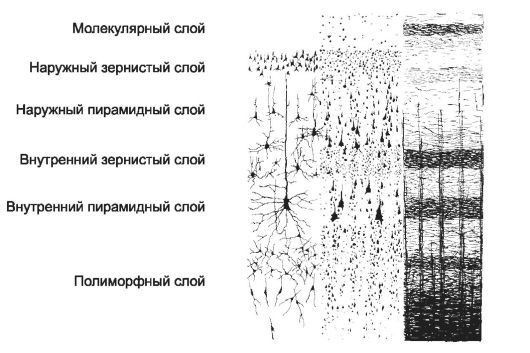 Riso. 8-39. Citoarchitettonica e mieloarchitettonica della corteccia cerebrale. Usando vari metodi colorazione, è possibile ottenere un'immagine della struttura della corteccia con processi neuronali ben definiti ( lato sinistro figura), solo perikarya (parte centrale della figura) o fibre mielinizzate (parte destra della figura). In quest'ultimo caso, a livello degli strati granulare interno e piramidale interno, sono visibili accumuli compatti di fibre mieliniche a forma di striscia rispettivamente della lamina granulare esterna e interna.
Riso. 8-39. Citoarchitettonica e mieloarchitettonica della corteccia cerebrale. Usando vari metodi colorazione, è possibile ottenere un'immagine della struttura della corteccia con processi neuronali ben definiti ( lato sinistro figura), solo perikarya (parte centrale della figura) o fibre mielinizzate (parte destra della figura). In quest'ultimo caso, a livello degli strati granulare interno e piramidale interno, sono visibili accumuli compatti di fibre mieliniche a forma di striscia rispettivamente della lamina granulare esterna e interna.
Neuroni corticali
neuroni piramidali. Le dimensioni del pericarion sono di 10-100 µm, hanno un lungo dendrite apicale che emerge dalla sommità della piramide, e altri dendriti che si estendono dalle superfici laterali del pericarion. Un assone parte dalla base della piramide, estendendosi nella sostanza bianca. I rami collaterali ricorrenti dell'assone terminano in corrispondenza di altri neuroni piramidali o neuroni corticali intercalari. Gigante neuroni piramidali situati nello strato V della corteccia motoria, questi sono i più grandi neuroni corticali. La dimensione del loro pericarion è superiore a 100 micron, danno origine a grandi assoni mielinizzati del tratto piramidale. Grandi neuroni piramidali si trovano anche nello strato V della corteccia visiva. Lobo occipitale. Inviano assoni al tronco cerebrale e sono coinvolti nel riflesso del movimento oculare.
sistema piramidale. Nel controllo cosciente dei movimenti volontari prende parte la corteccia dei lobi frontali del cervello (giro precentrale), da dove gli impulsi nervosi attraverso i motoneuroni del midollo spinale arrivano allo scheletro
muscoli di volo dell'arto. Il neurotrasmettitore più importante nella trasmissione degli impulsi tra i neuroni in questo sistema è l'acetilcolina. neuroni stellati. I loro perikarion sono arrotondati, poligonali o forma triangolare, 4-8 µm di diametro. L'assone e i dendriti si estendono a breve distanza dal pericarion e partecipano alla formazione delle connessioni intracorticali.
Neuroni fusiformi più comune nello strato VI. I dendriti derivano dalle estremità opposte del pericarion. L'assone si estende in profondità nella sostanza bianca. Neuroni piramidali invertiti presente in tutti gli strati tranne il primo. Hanno un pericarione poligonale e brevi dendriti. L'assone è diretto verticalmente alla superficie della corteccia, emettendo collaterali in tutti gli strati. Neuroni orizzontali sono nello strato I. Un lungo assone parte dal loro pericarione fusiforme, che, insieme ai dendriti, forma connessioni orizzontali all'interno dello strato I.
Moduli e cilindri della corteccia
Abbaio emisferi composto da moduli e cilindri.
Modulo ha un diametro di circa 0,1 mm e penetra per tutto lo spessore della corticale. Nella corteccia visiva, un modulo tipico include più di 100.000 cellule connesse sinapticamente che formano reti neurali. I moduli sono in contatto tra loro da collaterali di dendriti e assoni.
Cilindri. C'è un'idea di connessioni verticali (radali) da uno a trecento neuroni corticali, che insieme formano una certa unità funzionale chiamata cilindro. Una tale struttura può raggiungere diverse centinaia di micron di diametro. I cilindri sono tracciati più chiaramente nei campi sensoriali della corteccia, dove ricevono informazioni da determinati gruppi di neuroni afferenti.
GANGLI BASALI
I gangli della base formano un complesso di ammassi sottocorticali di neuroni situati nella sostanza bianca centrale degli emisferi cerebrali. Questi gangli forniscono la regolazione delle funzioni motorie e vegetative, partecipano all'attuazione di processi integrativi di attività nervosa superiore. I gangli della base si uniscono nucleo caudato (nucleo caudato), conchiglia (putamen) palla pallida (globo pallido) e materia nera (sostanza nera). I gangli della base ricevono input dalla corteccia frontale, che è responsabile del controllo dei movimenti volontari, e mediano il controllo del movimento involontario inverso attraverso la corteccia premotoria e il talamo.
nuclei motori i gangli della base sono divisi in diversi funzionalmente vari gruppi. Uno di questi è chiamato striato. Include il nucleo caudato e il putamen. Entrambe queste strutture striatali raccolgono informazioni da altre aree del cervello. I neuroni in quasi tutte le aree della neocorteccia inviano assoni allo striato. Il guscio termina con gli assoni
neuroni situati nella corteccia temporale, parietale e frontale, nonché nella parte compatta della substantia nigra.
sistema extrapiramidale. Per un processo di movimento olistico, è importante controllare non solo i movimenti volontari, ma anche quelli involontari. Questa funzione è svolta dal sistema extrapiramidale, che garantisce movimenti fluidi e la capacità di interrompere l'azione iniziata. Questo sistema unisce i gangli della base situati all'esterno midollo allungato fuori dalle piramidi (da cui il termine "sistema extrapiramidale"). Il controllo inconscio sui movimenti eseguiti da questo sistema è fornito dal neurotrasmettitore dopamina.
MEMBRI DEL CERVELLO
Il cervello e il midollo spinale sono protetti da membrane (Fig. 8-42). Pia madre (Pia madre) direttamente adiacente al cervello. Fuori dalla dura madre (dura madre). Tra di loro c'è la membrana aracnoidea media (t. aracnoidea). Tutti i gusci sono formati da tessuto connettivo fibroso. I gusci molli e aracnoidi possono essere considerati come un'unità sotto il nome generale pia-aracnoidea, O leptomeninge.
Morbido le meningi contengono fasci intrecciati di fibre di collagene e una rete di fibre elastiche, molti vasi sanguigni uniformemente distribuiti. All'esterno, il guscio è ricoperto da uno strato di cellule piatte. Il guscio ripete il corso dei solchi e delle circonvoluzioni del cervello.
ragnatela guscio - una rete di sottili partizioni di tessuto connettivo (trabecole), costituita principalmente da fibre di collagene e una piccola quantità
 Riso. 8-42. Meningi. 1 - cellula endoteliale; 2 - dura madre; 3 - guscio aracnoideo; 4 - tessuto connettivo; 5 - cellule dell'aracnoide; 6 - macrofago; 7 - spazio subaracnoideo; 8 - pia madre; 9 - vaso sanguigno; 10 - tessuto connettivo perivascolare; 11 - cervello.
Riso. 8-42. Meningi. 1 - cellula endoteliale; 2 - dura madre; 3 - guscio aracnoideo; 4 - tessuto connettivo; 5 - cellule dell'aracnoide; 6 - macrofago; 7 - spazio subaracnoideo; 8 - pia madre; 9 - vaso sanguigno; 10 - tessuto connettivo perivascolare; 11 - cervello.
Onori di fibre elastiche. La membrana aracnoidea è rivestita dall'interno e dall'esterno con uno strato continuo di sottili cellule appiattite. Lo spazio tra le trabecole è riempito di liquido cerebrospinale ed è chiamato spazio subaracnoideo. L'aracnoide nella regione dei solchi cerebrali non è strettamente adiacente alla pia madre. Ci sono cisterne contenenti una grande quantità di liquido cerebrospinale. solido le meningi sono costituite da tessuto connettivo fibroso denso dominato da fibre di collagene. Lo spazio tra la dura madre e l'aracnoide è chiamato spazio subdurale. Contiene una piccola quantità di liquido diverso dal liquido cerebrospinale. All'esterno, la dura madre è ricoperta da tessuto connettivo lasso grande quantità vene. La dura madre forma pieghe che, insieme al periostio, formano seni rivestiti di endotelio e contenenti sangue venoso. Lo spazio tra duro meningi e il periostio è chiamato epidurale. Contiene tessuto connettivo lasso e il plesso venoso vertebrale interno.
SENSORI
Gli organi di senso sono strutture sensoriali per la percezione di stimoli visivi, uditivi, olfattivi e gustativi. Di conseguenza, questi sono gli organi della vista, dell'udito e dell'equilibrio, dell'olfatto e del gusto.
Visione
L'organo della visione è costituito dal bulbo oculare collegato attraverso nervo ottico con il cervello, e comprende apparato ausiliario sotto forma di palpebre, ghiandola lacrimale e muscoli oculomotori striati.
La parete del bulbo oculare è formata da tre gusci: quello esterno fibroso (nella parte posteriore della sclera opaca, passando nella cornea trasparente nella parte anteriore; il limbus è il confine tra la cornea e la sclera), quello di mezzo è il vascolare vero e proprio (coroideo) e interno - mesh (Fig. 8-45).
Cornea(cornea)- guscio trasparente parete anteriore dell'occhio, consiste di cinque strati. Sulla superficie della cornea è presente un film di secrezioni delle ghiandole lacrimali e mucose, contenente lisozima, lattoferrina, immunoglobuline. La superficie anteriore della cornea è epitelio squamoso stratificato non cheratinizzato. Strato omogeneo privo di cellule della sostanza fondamentale e fibre sottili di collagene e reticolina orientate in modo casuale - membrana limitante anteriore- mantiene la forma della cornea. propria sostanza rappresentato da placche di collagene correttamente posizionate e fibroblasti appiattiti immersi nel tappeto
 Riso. 8-45. Parte anteriore dell'occhio.
Riso. 8-45. Parte anteriore dell'occhio.
rici dai glicosaminoglicani. Membrana limitante posteriore- uno strato trasparente della cornea, situato tra la propria sostanza e l'endotelio della superficie posteriore della cornea, costituito da fibre di collagene di tipo VIII e una sostanza amorfa. Endotelio corneale delimita la camera anteriore dell'occhio di fronte, questo è uno strato di cellule praticamente non proliferanti che svolge le funzioni di barriera e di "pompaggio" necessarie per l'idratazione dello stroma corneale e per mantenerne la trasparenza.
Sclera- guscio opaco esterno del bulbo oculare. La sclera è costituita da fili densi di fibre di collagene, tra i quali vi sono fibroblasti appiattiti. Alla giunzione della sclera con la cornea, ci sono piccole cavità comunicanti, che insieme si formano seno venoso della sclera(seno venoso sclerale), fornendo un deflusso di fluido dalla camera anteriore dell'occhio. coroide fornisce nutrimento alla retina. Questa membrana è costituita da diverse placche (strati): sopravascolare, vascolare, coriocapillare e basale.
placca sopravascolare(lamina suprachoroidea) situato al confine con la sclera, è costituito da tessuto connettivo fibroso lasso con numerose cellule pigmentate.
Placca vascolare(lamina vasculosa) contiene un plesso di arterie e vene; nel tessuto connettivo lasso si trovano le cellule del pigmento e le SMC.
Capillare vascolare(lamina coroidocapillare)piatto formato da un plesso di capillari sinusoidali.
Situato al confine con la retina lamina basale(lamina basale)(membrana basale e fibre elastiche). Davanti all'occhio, la coroide forma l'iride e il corpo ciliare.
iris(iride) - una continuazione della coroide, situata tra la cornea e il cristallino, separa le camere anteriore e posteriore dell'occhio e contiene l'endotelio, il confine esterno, il confine vascolare, il confine interno e gli strati del pigmento posteriore. Endotelio(epitelio anteriore) - continuazione dell'endotelio corneale. Esterno E confine interno gli strati hanno una struttura simile e contengono fibroblasti e melanociti immersi nella sostanza fondamentale. Vascolare strato - tessuto connettivo lasso contenente numerosi vasi e melanociti. Lo strato di pigmento posteriore passa in un epitelio retinico a due strati che copre il corpo ciliare. Come parte dell'iride, si formano gli HMC pupilla dello sfintere(M. sfintere pupillare) E dilatatore della pupilla(M. dilatator pupillae). Quando le fibre nervose parasimpatiche (colinergiche) vengono stimolate, la pupilla si restringe, la stimolazione simpatica porta alla dilatazione della pupilla. Corpo ciglia. Nella regione dell'angolo dell'occhio, la coroide si ispessisce, formando il corpo ciliare. (corpo ciliare), avente sul taglio la forma di un triangolo, rivolto dalla base nella camera anteriore. La massa principale del corpo ciliare è occupata dal muscolo ciliare (m. ciliaris). Nella sua composizione, gli MMC corrono in tre direzioni reciprocamente perpendicolari. Dal corpo ciliare si dipartono verso il cristallino processi ciliari(processo ciliare). Contengono una massa di capillari, ricoperti da due strati di epitelio: pigmento e secretorio, che producono umore acqueo. Attaccato ai processi ciliari fascia per ciglia(zonula ciliare). Le fibre colinergiche postgangliari del ganglio ciliare terminano nel muscolo ciliare. Quando si riduce muscolo ciliare la fascia ciliare si rilassa e il rigonfiamento del cristallino aumenta.
lente(lente) ha la forma di una lente biconvessa. La sua superficie anteriore è ricoperta da un singolo strato di epitelio cuboidale, che diventa più alto verso l'equatore. cellule epiteliali lente sono collegati da giunzioni gap. Trasparente fibre della lente(fibre lentis) costituiscono la parte principale del cristallino e contengono cristallini. capsula del cristallino(capsula lentis)- spessa membrana basale con un contenuto significativo di fibre di reticolina.
Fotocamera frontaleè delimitato anteriormente dalla cornea, posteriormente dall'iride e all'interno della pupilla dalla parte centrale della superficie anteriore del cristallino. La profondità della camera anteriore è massima nella parte centrale, dove raggiunge i 3-3,5 mm. Angolo tra superficie posteriore parte periferica della cornea e la superficie anteriore della radice dell'iride - angolo della camera anteriore si trova nella regione di transizione della cornea alla sclera e dell'iride al corpo ciliare.
videocamera posteriore- lo spazio dietro l'iride, delimitato dal cristallino, corpo ciliare e corpo vitreo.
umore acqueo(umorismo aquosus)- fluido contenente proteine plasmatiche solubili e depolimerizzate acido ialuronico. L'umidità acquosa non contiene fibrinogeno ed è ipertonica rispetto al sangue. La maggior parte umore acqueo proviene dai capillari e dall'epitelio dei processi ciliari in videocamera posteriore e passa tra l'iride e il cristallino nella camera anteriore. Rete trabecolare(reticolo trabecolare)È formato da elementi della cornea, dell'iride e del corpo ciliare ed è importante per il deflusso dell'umor acqueo dalla camera anteriore dell'occhio.
Pressione intraoculare(23 mm Hg) dipende dall'equilibrio tra la quantità di umore acqueo formato e rimosso; la velocità della sua formazione e assorbimento è di circa 2 mm 3 /min. Principale via di deflusso dell'umor acqueo: trabecolato → seno venoso della sclera → vasi venosi occhi.
Barriere ematotessutali regolare lo scambio tra sangue e tessuti dell'occhio. Lo scambio tra sangue e umor acqueo avviene nell'iride e nel corpo ciliare. Le cellule dell'epitelio ciliare sono collegate da giunzioni strette che limitano il trasporto di macromolecole. I capillari dell'iride non hanno fenestrae.
corpo vitreo(corpo vitreo)- mezzo trasparente dell'occhio, riempie la cavità tra il cristallino e la retina. Il corpo vitreo è un gel contenente acqua, collagene di tipo II, IX e XI, proteina vitreina e acido ialuronico. Il canale vitreo passa attraverso il corpo vitreo dalla retina al cristallino (canalis hyaloidus)- residuo embrionale sistema vascolare occhi. membrana vetrosa (membrana vitrea)- un accumulo di fibre di collagene lungo la periferia del corpo vitreo, come se ne formassero la capsula.
Retina
retina (retina, retina)- guscio interno occhi, ha reparto visivo, lungo il bordo frastagliato (ora serrata) passando nella sezione cieca, coprendo la parte posteriore del corpo ciliare e l'iride. Sul bordo posteriore dell'asse ottico dell'occhio, la retina ha una forma arrotondata macchia gialla(macula lutea) circa 2 mm di diametro. Fossa centrale(fovea centrale)- recesso nel mezzo macchia gialla, il luogo della migliore percezione. nervo ottico(nervo ottico) esce dalla retina medialmente alla macula. Qui si forma disco ottico(discus nervi ottici). Al centro del disco è presente un incavo in cui sono visibili i vasi che irrorano la retina.
Strati retinici. La parte visiva della retina è formata in sequenza (dalla superficie esterna del bulbo oculare) da strati disposti (Fig. 8-46): pigmento, nucleare esterno, reticolare esterno, nucleare interno, reticolare interno e gangliare.
strato di pigmento(strato pigmentoso). Celle poligonali adiacenti a coroide. Una cella epitelio pigmentato interagisce
 Riso. 8-46. Retina. Le frecce a sinistra indicano le direzioni del flusso luminoso e dell'eccitazione. L'epitelio pigmentato circonda i segmenti esterni delle cellule dei fotorecettori che formano contatti sinaptici con i neuroni bipolari. Le informazioni dalle cellule bipolari vengono trasmesse alle cellule gangliari e vanno al cervello lungo i loro assoni, che formano il nervo ottico. Gli spazi tra i neuroni sono pieni di grandi cellule della glia radiale. I loro processi esterni terminano al confine tra i segmenti esterno e interno delle cellule dei fotorecettori.
Riso. 8-46. Retina. Le frecce a sinistra indicano le direzioni del flusso luminoso e dell'eccitazione. L'epitelio pigmentato circonda i segmenti esterni delle cellule dei fotorecettori che formano contatti sinaptici con i neuroni bipolari. Le informazioni dalle cellule bipolari vengono trasmesse alle cellule gangliari e vanno al cervello lungo i loro assoni, che formano il nervo ottico. Gli spazi tra i neuroni sono pieni di grandi cellule della glia radiale. I loro processi esterni terminano al confine tra i segmenti esterno e interno delle cellule dei fotorecettori.
con segmenti esterni di dozzine di cellule fotorecettrici - bastoncelli e coni. Le cellule epiteliali del pigmento immagazzinano la vitamina A, partecipano alla sua trasformazione e trasferiscono i suoi derivati alle cellule dei fotorecettori per la formazione pigmento visivo.
Nucleare all'aperto strato (strato nucleare esterno) comprende le parti nucleate delle cellule dei fotorecettori. coni concentrato nella macula. Il bulbo oculare è organizzato in modo tale che la parte centrale del punto luminoso dell'oggetto visualizzato cada sui coni. Alla periferia della macchia gialla si trovano bastoni.
Maglia esterna(strato plessiforme esterno). Qui vengono realizzati i contatti dei segmenti interni di bastoncelli e coni con i dendriti delle cellule bipolari.
nucleare interno(strato nucleare interno). Contiene cellule bipolari che collegano bastoncelli e coni alle cellule gangliari, nonché cellule orizzontali e amacrine. I perikarion delle cellule amacrine si trovano nella parte interna dello strato nucleare interno.
Rete interna(strato plessiforme interno). In esso, le cellule bipolari sono in contatto con le cellule gangliari, le cellule amacrine agiscono come
neuroni intercalari. Un concetto popolare è che un numero limitato di cellule bipolari trasmette informazioni a 16 tipi di cellule gangliari con la partecipazione di almeno 20 tipi di cellule amacrine.
Strato gangliare(strato gangliare) contiene neuroni gangliari. Schema generale la trasmissione dell'informazione nella retina è la seguente: cellula recettrice → cellula bipolare → cellula gangliare e contemporaneamente cellula amacrina → cellula gangliare.
cellule fotorecettrici- aste (Fig. 8-47) e coni. I processi periferici delle cellule dei fotorecettori sono costituiti dall'esterno e segmenti interni collegati da un ciglio. Distinguere tra centrale e visione periferica, che è associato alla natura della distribuzione di bastoncelli e coni nella retina. La fovea contiene prevalentemente coni. Ogni cono della fovea forma una sinapsi con un solo neurone bipolare. La visione centrale, così come l'acuità visiva, sono realizzate dai coni. La visione periferica, così come la visione notturna e la percezione degli oggetti in movimento, sono le funzioni delle aste.
Segmento esterno ha molti dischi chiusi appiattiti contenenti pigmenti visivi: rodopsina - in bastoncini; i pigmenti rosso, verde e blu sono nei coni.
 Riso. 8-47. Diagramma della bacchetta. Il segmento esterno contiene una pila di dischi contenenti il pigmento visivo rodopsina. La membrana del disco e la membrana cellulare sono separate. La luce (h) attiva la rodopsina Rh* nei dischi.
Riso. 8-47. Diagramma della bacchetta. Il segmento esterno contiene una pila di dischi contenenti il pigmento visivo rodopsina. La membrana del disco e la membrana cellulare sono separate. La luce (h) attiva la rodopsina Rh* nei dischi.
Segmento interno pieno di mitocondri e contiene un corpo basale, dal quale si estendono 9 paia di microtubuli nel segmento esterno.
Percezione del colore- funzione dei coni. Esistono tre tipi di coni, ciascuno contenente solo uno dei tre diversi pigmenti visivi (rosso, verde e blu).
pigmento visivo costituito da apoproteine (opsina), legato covalentemente al cromoforo (11-cis-retinale o 11-NMC-deidroretinico).
neuroni retinici(cellule bipolari, orizzontali, amacrine e gangliari) sintetizzano acetilcolina, dopamina, acido L-glutammico, glicina, acido γ-amminobutirrico. Alcuni neuroni contengono serotonina, suoi analoghi (indolamine) e neuropeptidi.
Celle orizzontali. I pericarioni si trovano nella parte esterna dello strato nucleare interno e i processi entrano nella regione della sinapsi tra i fotorecettori e le cellule bipolari. Le celle orizzontali ricevono informazioni dai coni e le trasmettono anche ai coni.
cellule amacrine. I loro perikarya si trovano nella parte interna dello strato nucleare interno nell'area delle sinapsi tra le cellule bipolari e gangliari.
cellule bipolari rispondere al contrasto dell'immagine. Alcuni bipolari rispondono più fortemente al colore che al contrasto in bianco e nero. Alcuni ricevono informazioni principalmente dai bastoncelli, altri dai coni.
cellule gangliari - grandi neuroni multipolari di molte varietà. I loro assoni formano il nervo ottico. Le cellule gangliari rispondono a molte proprietà dell'oggetto visivo (ad esempio, oggetti chiari e scuri, uniformità dell'illuminazione, colore dell'oggetto, suo orientamento).
Glia della retina. Oltre ai neuroni, la retina contiene grandi cellule gliali radiali (vedi Figura 8-46). I loro nuclei si trovano nello strato nucleare interno. I processi esterni terminano in microvilli, formando lo strato limite esterno. I processi interni hanno un'estensione (peduncolo) nello strato limite interno al confine con il corpo vitreo. Le cellule gliali svolgono un ruolo importante nella regolazione dell'omeostasi degli ioni retinici.
Ghiandola lacrimale
La ghiandola lacrimale è formata da un gruppo di complesse ghiandole tubolare-alveolari; le loro sezioni secretorie sono circondate da cellule mioepiteliali. Il segreto della ghiandola, il liquido lacrimale, attraverso 6-12 dotti entra nel fornice della congiuntiva e lava la cornea e la congiuntiva.
Odore
La parte periferica dell'analizzatore olfattivo (Fig. 8-49) è rappresentata dal rivestimento olfattivo (area olfattiva), che occupa la parte mediana della conca nasale superiore e la corrispondente sezione della mucosa del setto nasale con un'area di 2,5 cm2. Ob-
epitelio giovanile contiene cellule recettrici. il cui numero totale raggiunge i 50 milioni I loro processi centrali (assoni) trasmettono informazioni al bulbo olfattivo. Le cellule del recettore olfattivo sono circondate da cellule di supporto. L'epitelio olfattivo contiene anche cellule del pigmento, su cui giallo rivestimento olfattivo. Nel tessuto connettivo subepiteliale si trovano le sezioni terminali delle ghiandole di Bowman, vasi sanguigni e fasci di fibre non mielinizzate del nervo olfattivo. Il muco secreto dalle ghiandole di Bowman ricopre la superficie del rivestimento olfattivo con uno strato spesso 60 µm. Le ciglia olfattive immerse nel muco sono coinvolte nel processo di chemosensing. Nervo olfattivo- un insieme di sottili filamenti olfattivi che passano attraverso i fori dell'osso etmoide fino ai bulbi olfattivi e formano fasci di 10-100 filamenti.
cellula olfattiva
Morfologia. Il corpo della cellula olfattiva contiene numerosi mitocondri, cisterne del reticolo endoplasmatico con ribosomi, elementi del complesso del Golgi e lisosomi. Le cellule olfattive, oltre a quella centrale (assone), hanno un breve processo periferico (dendrite), che termina sulla superficie dell'epitelio olfattivo con un ispessimento sferico - un club olfattivo con un diametro di 1-2 micron. Contiene mitocondri, piccoli vacuoli e corpi basali per 20-80 peli olfattivi lunghi 30-200 µm che si estendono dalla sommità della clava, aventi la struttura delle tipiche ciglia. I recettori degli odori sono presenti nella membrana delle ciglia olfattive.
Funzione. Le cellule recettrici del rivestimento olfattivo registrano 25-35 odori primari, ma le loro combinazioni formano molti milioni di odori percepiti. I neuroni del rivestimento olfattivo della cavità nasale umana hanno 350 diversi recettori olfattivi. Ogni neurone del rivestimento olfattivo esprime un solo gene del recettore dell'odore, ma ogni recettore riconosce più odori. Pertanto, un particolare odore dipende dall'attivazione di una certa combinazione di recettori. Rigenerazione. La durata della vita delle cellule olfattive è di 40 giorni. Le cellule del recettore olfattivo sono un'eccezione tra i neuroni: sono costantemente aggiornate grazie alle cellule precursori, ad es. appartengono a una popolazione cellulare in rinnovamento. I precursori delle cellule recettrici sono le cellule basali dell'epitelio del rivestimento olfattivo.
Gusto
La parte periferica dell'analizzatore del gusto sono le papille gustative (Fig. 8-51). Si trovano nell'epitelio della bocca, della lingua, della faringe anteriore
 Riso. 8-49. Organo olfattivo. I neuroni del recettore olfattivo si trovano nell'epitelio del rivestimento olfattivo. I loro processi esterni terminano in club olfattivi, da cui lati diversi I peli olfattivi si estendono parallelamente alla superficie dell'epitelio. Assoni dei neuroni recettori nei filamenti olfattivi (fila olfattiva) passaggio del nervo olfattivo lamina cribrosa etmoide e formano sinapsi con i neuroni del bulbo olfattivo (bulbo olfattivo). Numerose cellule di supporto circondano le cellule recettrici.
Riso. 8-49. Organo olfattivo. I neuroni del recettore olfattivo si trovano nell'epitelio del rivestimento olfattivo. I loro processi esterni terminano in club olfattivi, da cui lati diversi I peli olfattivi si estendono parallelamente alla superficie dell'epitelio. Assoni dei neuroni recettori nei filamenti olfattivi (fila olfattiva) passaggio del nervo olfattivo lamina cribrosa etmoide e formano sinapsi con i neuroni del bulbo olfattivo (bulbo olfattivo). Numerose cellule di supporto circondano le cellule recettrici.
ki, esofago e laringe. La loro localizzazione principale sono le papille chemiosensibili della lingua: a forma di fungo, scanalate ea forma di foglia. Nei bambini e meno spesso negli adulti, le papille gustative si trovano nell'epitelio delle labbra, nell'epiglottide e persino nelle corde vocali.
papilla gustativa
papilla gustativa (gemma gustatoria) ha forma ellissoidale, altezza 27-115 micron e larghezza 16-70 micron. Nella sua sezione apicale è pieno di sostanza amorfa canale del gusto, apertura sulla superficie dell'epitelio con un poro gusto. La papilla gustativa è formata da 30-80 cellule allungate, strettamente adiacenti l'una all'altra. La maggior parte di queste cellule entra in contatto con le fibre nervose che penetrano nel rene dal plesso nervoso subepiteliale. Le cellule delle papille gustative sono morfologicamente eterogenee. Ci sono quattro tipi di cellule.

Riso. 8-51. papilla gustativa consiste di celle leggere allungate vari tipi. La loro superficie apicale si apre nel poro del gusto sulla superficie dello strato epiteliale. Le cellule delle papille gustative formano sinapsi con i processi periferici dei neuroni sensoriali.
cellule chemocettrici. Sebbene tutti i tipi di cellule formino contatti con fibre afferenti, la funzione di chemosensing è associata alle cellule di tipo III. Nella regione presinaptica delle cellule gustative, le vescicole granulari contengono serotonina, un neurotrasmettitore sinaptico afferente. Le cellule chemocettrici percepiscono il dolce, l'acido, l'amaro, il salato e il gusto del glutammato (umami, umami). Rigenerazione. Nella papilla gustativa c'è un costante rinnovamento delle cellule. Dalla regione periferica della papilla gustativa, le cellule si spostano verso la sua parte centrale ad una velocità di 0,06 µm/ora. Durata media vita delle cellule gustative organo recettoreè di 250 ± 50 ore.
Udito ed equilibrio
Organo dell'UDITO
orecchio esterno include padiglione auricolare, all'aperto canale uditivo E timpano trasmissione vibrazioni sonore sugli ossicini dell'orecchio medio. Il padiglione auricolare è formato da cartilagine elastica ricoperta di pelle sottile. Il meato uditivo esterno è rivestito di pelle contenente follicoli piliferi, tipico ghiandole sebacee e ghiandole ceruminose - ghiandole sebacee modificate che producono cerume. La superficie esterna del timpano è ricoperta di pelle. Dall'interno, dal lato della cavità timpanica (orecchio medio), la membrana timpanica (vedi Fig. 8-62) è rivestita da un epitelio cuboidale monostrato situato su una sottile lamina di tessuto connettivo.
Orecchio medio contiene gli ossicini uditivi - martello, incudine e staffa, che trasmettono le vibrazioni dal timpano alla membrana finestra ovale(Vedi Figura 8-62). La cavità timpanica è rivestita da epitelio stratificato, che si trasforma in un ciliato cilindrico monostrato all'apertura del tubo uditivo. L'osso della parete mediale della cavità timpanica presenta due finestre, ovale e rotonda, che si separano cavità timpanica dal labirinto osseo dell'orecchio interno.
orecchio interno formato da labirinto osseo osso temporale, che contiene un labirinto membranoso che ne ripete il rilievo (Fig. 8-62, Fig. 8-63). Labirinto osseo- il sistema di canali semicircolari e la cavità comunicante con essi - il vestibolo (vestibolo).labirinto membranoso- un sistema di tubi e sacche di tessuto connettivo a parete sottile situati all'interno labirinto osseo. Nelle ampolle ossee, i canali membranosi si espandono. Nel vestibolo, il labirinto membranoso forma due sacche interconnesse: borsa ellittica(grembo, utricolo), in cui si aprono i canali semicircolari membranosi, e tasca sferica(borsa, sacculo). I canali semicircolari membranosi e le sacche del vestibolo sono riempiti di endolinfa e comunicano con la coclea, nonché con il sacco endolinfatico situato nella cavità cranica, dove l'endolinfa viene riassorbita. Il rivestimento epiteliale del sacco endolinfatico contiene cellule cilindriche con citoplasma denso e nuclei. forma irregolare, così come cellule cilindriche con citoplasma leggero, microvilli alti, numerose vescicole pinocitiche e vacuoli.
Lumaca
Lumaca (coclea)- canale osseo attorcigliato a spirale. La coclea forma 2,5 spirali lunghe circa 35 mm. Le membrane basilare (principale) e vestibolare situate all'interno del canale cocleare dividono la sua cavità in tre parti, questa scala a tamburo(scala timpanica),scala del vestibolo(scala vestiboli) E canale membranoso lumache (scala media, scala mediana, passaggio cocleare) (Fig. 8-56). L'endolinfa riempie il canale membranoso della coclea e la perilinfa riempie la scala vestibolo e la scala timpanica. La scala vestibolo e la scala timpanica comunicano all'apice della coclea attraverso un forame (elicotrema). Nel canale membranoso della coclea si trova la membrana basilare apparato recettore lumache - organo a spirale.
Endolinfa- un liquido viscoso che riempie il canale membranoso della coclea e si collega attraverso un canale speciale (dotto ricongiunto) con endolinfa apparato vestibolare. La concentrazione di K+ nell'endolinfa è 100 volte maggiore che nel liquido cerebrospinale e nella perilinfa; La concentrazione di Na+ nell'endolinfa è 10 volte inferiore a quella della perilinfa.
Perilinfa Di Composizione chimica vicino al plasma sanguigno liquido cerebrospinale e occupa una posizione intermedia tra loro in termini di contenuto proteico.
organo a spirale
L'organo a spirale (Fig. 8-57) contiene diverse file di cellule ciliate a contatto con la membrana tegumentaria. Ci sono capelli interni ed esterni e cellule di supporto. Le cellule ciliate interne sono recettori, formano contatti sinaptici con i processi periferici dei neuroni sensoriali del ganglio spirale.
cellule ciliate interne formare una riga. Le cellule sul lato laterale sono ricoperte da cellule pilastro interne e con le loro altre superfici sono in contatto con cellule falangee. Nella parte apicale sono presenti stereociglia passanti per la cuticola, costituite da 20-50 microvilli immobili. Le stereociglia si trovano a semicerchio (oa forma di lettera V), aperte verso le strutture esterne dell'organo a spirale. Le cellule ciliate interne sono eccitate in risposta a stimolo sonoro e trasmettono l'eccitazione alle fibre afferenti nervo uditivo. Lo spostamento della membrana tegumentaria provoca la deformazione della stereocilia, nella cui membrana si aprono i canali ionici meccanosensibili e si verifica la depolarizzazione della membrana cellulare.
cellule ciliate esterne altezza da 20 a 70 micron disposti su 3-5 file, hanno forma cilindrica e stereociglia. La parte basale si trova in una depressione formata dalle cellule falangee esterne. La superficie laterale delle celle viene lavata da un liquido che riempie il tunnel esterno (cunicolo esterno) e supporto
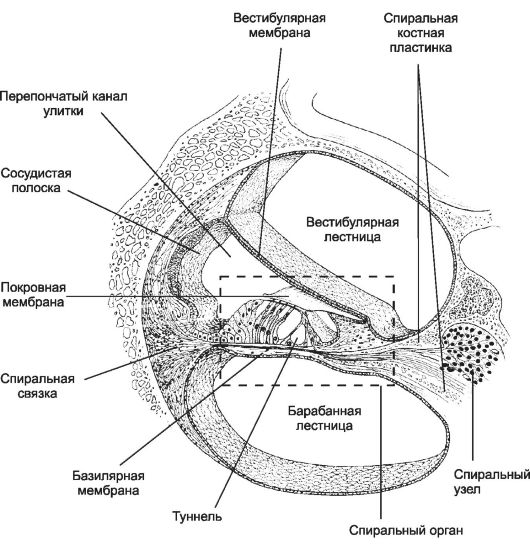 Riso. 8-56. canale membranoso e organo a spirale. Il canale cocleare è diviso in scala vestibuli, scala tympani e canale membranoso, in cui si trova l'organo a spirale. Il canale membranoso è separato dalla scala timpanica dalla membrana basilare. Contiene processi periferici di neuroni del ganglio spirale, che formano contatti sinaptici con le cellule ciliate esterne ed interne.
Riso. 8-56. canale membranoso e organo a spirale. Il canale cocleare è diviso in scala vestibuli, scala tympani e canale membranoso, in cui si trova l'organo a spirale. Il canale membranoso è separato dalla scala timpanica dalla membrana basilare. Contiene processi periferici di neuroni del ganglio spirale, che formano contatti sinaptici con le cellule ciliate esterne ed interne.
peloso spazio cellulare (Cuniculus intermedio). Le cellule ciliate esterne formano sinapsi con le fibre efferenti del tratto olivococleare. La secrezione di acetilcolina provoca l'iperpolarizzazione delle cellule ciliate esterne, che ha un effetto modulante sull'altezza delle cellule e le successive reazioni nella catena di trasmissione della stimolazione sonora.
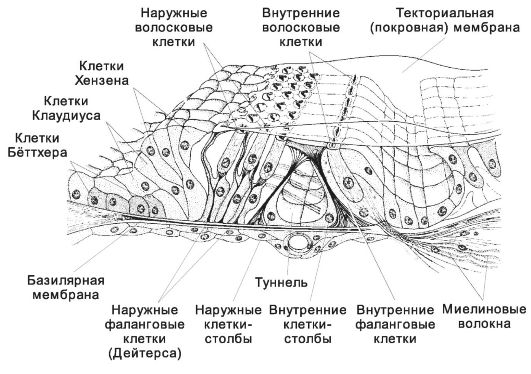 Riso. 8-57. organo a spirale. Le cellule ciliate meccanosensibili formano diverse file: una fila interna e 3-5 file esterne. Le cellule ciliate interne ed esterne sono separate da un tunnel. È formato da grandi cellule pilastro esterne ed interne. La membrana tegumentaria è in contatto con la stereocilia delle cellule ciliate.
Riso. 8-57. organo a spirale. Le cellule ciliate meccanosensibili formano diverse file: una fila interna e 3-5 file esterne. Le cellule ciliate interne ed esterne sono separate da un tunnel. È formato da grandi cellule pilastro esterne ed interne. La membrana tegumentaria è in contatto con la stereocilia delle cellule ciliate.
cellule di supporto. Tra le cellule di supporto (supporto), ci sono cellule falangee interne, cellule falangee esterne, cellule colonnari del bordo esterno, cellule colonnari di supporto esterne, cellule cuboidali di supporto esterne, cellule ghiandolari basali esterne. Le cellule falangee entrano in contatto con le cellule ciliate, situate sulla membrana basilare. I processi delle cellule falangee esterne corrono paralleli alle cellule ciliate esterne, non toccandole per una distanza considerevole, ma a livello della parte apicale delle cellule ciliate entrano in contatto con esse. Le celle di supporto sono collegate da giunzioni gap. Le giunzioni gap sono coinvolte nel ripristino del livello di K + nell'endolinfa durante le reazioni in tracce dopo l'eccitazione delle cellule ciliate.
Membrana tegumentaria(membrana tectoria)È costituito da proteine (principalmente collagene), carboidrati e glicosaminoglicani, che formano la matrice e le fibre.
Membrana basilare presentato in più parti. Parte centrale - zona arcuata (pars tecta), parte laterale - zona pettine (parte pectinata). La zona arcuata è parzialmente incorporata nella placca a spirale ossea. Pertanto, è meno mobile quando si spostano fluidi cocleari in risposta alla stimolazione sonora. Al contrario, la zona pettinea è più mobile e vibra in risposta ai segnali sonori. Via di trasmissione dell'irritazione uditiva. Catena di trasmissione pressione sonora sembra nel seguente modo: membrana timpanica → martello → incudine → staffa → membrana della finestra vestibolare → perilinfa → membrane basilare e tegumentaria - membrana della finestra cocleare (Fig. 8-62). Quando la staffa viene spostata, le particelle perilinfatiche si muovono lungo la scala vestibuli e quindi attraverso l'elicotrema lungo la scala timpanica fino alla finestra rotonda. Il fluido spostato dallo spostamento della membrana della finestra vestibolare crea una pressione eccessiva nel canale vestibolare. Sotto l'influenza di questa pressione, la sezione basale della membrana principale viene spostata verso la scala timpanica. Una reazione oscillatoria sotto forma di onda si propaga dalla parte basale della membrana principale all'elicotrema. Lo spostamento della membrana tegumentaria rispetto alle cellule ciliate sotto l'azione del suono provoca la loro eccitazione. Lo spostamento della membrana rispetto all'epitelio sensoriale devia la stereocilia delle cellule ciliate, che apre i canali meccanosensibili nella membrana cellulare e porta alla depolarizzazione cellulare. La reazione elettrica risultante (microfono effetto) nella sua forma ripete la forma del segnale audio.
CORPO EQUILIBRIO
Ogni dilatazione ampulare del canale semicircolare contiene creste, O scaloppine(Fig. 8-62, 8-63). Le aree sensibili nelle sacche sono chiamate patch.
La composizione dell'epitelio di macchie e creste include sensibile peloso E cellule di supporto(Figura 8-64). Le cellule ciliate formano gruppi di diverse centinaia di unità. All'interno di ciascun gruppo, le cinocilia sono orientate allo stesso modo, ma il loro orientamento differisce tra i gruppi. L'epitelio delle macchie è ricoperto da una membrana otolitica gelatinosa. Otoliti- cristalli di carbonato di calcio. L'epitelio delle creste è circondato da una cupola gelatinosa trasparente.
cellule ciliate(Fig. 8-65) sono presenti in ciascuna ampolla dei canali semicircolari e nelle macchie delle sacche del vestibolo. Esistono due tipi di cellule ciliate. Le cellule di entrambi i tipi nella parte apicale contengono 40-110 peli immobili (stereocilia) e un ciglio (kinocilium) situato alla periferia del fascio di stereocilia. Le stereocilia più lunghe si trovano vicino al cinocilio, mentre la lunghezza del resto diminuisce con la distanza dal cinocilio. Cellule di tipo I situate al centro delle capesante, hanno la forma di un'anfora con fondo arrotondato e sono poste nella cavità a calice della terminazione nervosa afferente. Le fibre efferenti formano terminazioni sinaptiche sulle fibre afferenti associate alle cellule di tipo I.
 Riso. 8-62. Organo vestibolococleare. La pressione sonora attraverso la membrana timpanica lungo la catena ossea dell'orecchio medio (martello, incudine, staffa) viene trasmessa alla membrana della finestra del vestibolo e successivamente alla perilinfa delle scale che comunicano attraverso l'elicotrema alla finestra cocleare. Le vibrazioni della perilinfa vengono trasmesse alla membrana tegumentaria. Il suo spostamento provoca irritazione delle cellule ciliate.
Riso. 8-62. Organo vestibolococleare. La pressione sonora attraverso la membrana timpanica lungo la catena ossea dell'orecchio medio (martello, incudine, staffa) viene trasmessa alla membrana della finestra del vestibolo e successivamente alla perilinfa delle scale che comunicano attraverso l'elicotrema alla finestra cocleare. Le vibrazioni della perilinfa vengono trasmesse alla membrana tegumentaria. Il suo spostamento provoca irritazione delle cellule ciliate.
 Riso. 8-63. Aree neurosensoriali dell'orecchio interno evidenziato in nero. Le cellule ciliate meccanosensibili si trovano nelle aree neurosensoriali. Questi sono l'organo a spirale (organo dell'udito), creste e macchie o macula (organo dell'equilibrio).
Riso. 8-63. Aree neurosensoriali dell'orecchio interno evidenziato in nero. Le cellule ciliate meccanosensibili si trovano nelle aree neurosensoriali. Questi sono l'organo a spirale (organo dell'udito), creste e macchie o macula (organo dell'equilibrio).
![]() Riso. 8-64. Pettine. L'epitelio neurosensoriale è formato da capelli e cellule di supporto. Al centro del pettine ci sono cellule ciliate di tipo I e alla periferia di tipo II. Le cellule ciliate formano sinapsi con terminazioni nervose. Non ci sono otoliti nella cupola del pettine.
Riso. 8-64. Pettine. L'epitelio neurosensoriale è formato da capelli e cellule di supporto. Al centro del pettine ci sono cellule ciliate di tipo I e alla periferia di tipo II. Le cellule ciliate formano sinapsi con terminazioni nervose. Non ci sono otoliti nella cupola del pettine.
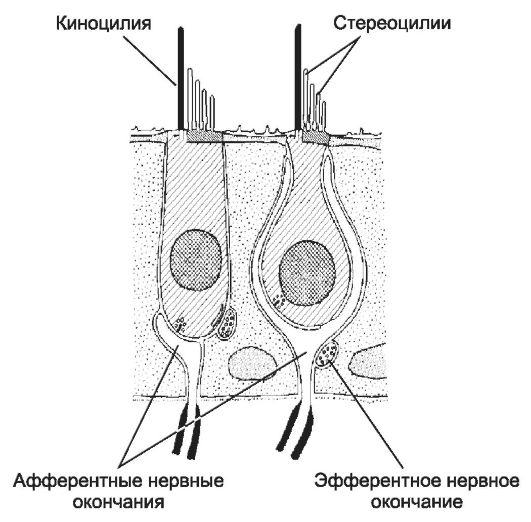 Riso. 8-65. cellule ciliate. A destra, una cella di tipo I; a sinistra, una cellula di tipo II.
Riso. 8-65. cellule ciliate. A destra, una cella di tipo I; a sinistra, una cellula di tipo II.
Cellule di tipo II situate lungo la periferia delle capesante, hanno l'aspetto di cilindri a base arrotondata. Le terminazioni nervose sulle cellule di tipo II possono essere afferenti (la maggior parte) o efferenti. Sensibilità direzionale Le cellule ciliate sono sensibili alla direzione dello stimolo. Quando lo stimolo è diretto dalla stereocilia al chinocilio, la cellula ciliata è eccitata. Con la direzione opposta dello stimolo, la risposta viene soppressa.
nervo vestibolare formato da processi di neuroni bipolari nel ganglio vestibolare. I processi periferici di questi neuroni si avvicinano alle cellule ciliate di ciascun canale semicircolare, utero e sacco, e quelli centrali vanno ai nuclei vestibolari del midollo allungato.


