Gli antipiretici per i bambini sono prescritti da un pediatra. Ma ci sono situazioni di emergenza per la febbre in cui il bambino deve ricevere immediatamente le medicine. Quindi i genitori si assumono la responsabilità e usano farmaci antipiretici. Cosa è permesso dare ai bambini? Come abbassare la temperatura nei bambini più grandi? Quali farmaci sono i più sicuri?
Conferenza per medici "Il ruolo degli ormoni nella regolazione del ciclo mestruale". Un corso di lezioni sull'ostetricia patologica per studenti universitari di medicina. conferenza per i medici Dyakova S.M., ostetrico-ginecologo, insegnante, esperienza lavorativa totale 47 anni.
Il ruolo degli ormoni nella regolazione del ciclo mestruale. Parte 1.
Il ruolo degli ormoni nella regolazione del ciclo mestruale. Parte 2.
Il ruolo degli ormoni nella regolazione del ciclo mestruale. Parte 3
Il ciclo mestruale e la sua regolazione
Il sistema riproduttivo (RS) svolge molte funzioni, la più importante delle quali è la continuazione della specie biologica. Attività funzionale ottimale sistema riproduttivo raggiunge i 16-18 anni, quando il corpo è pronto per concepire, partorire e nutrire un bambino. Una caratteristica della RS è anche il graduale sbiadimento varie funzioni: all'età di 45 anni, la funzione generativa svanisce, all'età di 50 anni - funzioni mestruali, quindi - ormonali.
Il sistema riproduttivo è costituito da cinque livelli: extrahypathalamic (corteccia cerebrale), ipotalamo, ipofisi, ovaie e organi e tessuti bersaglio (Fig. 1).
Il sistema riproduttivo funziona su base gerarchica, cioè il livello sottostante è subordinato a quello sovrastante (per i collegamenti diretti tra i legami di regolazione). La base della regolazione delle funzioni RS è il principio del feedback negativo tra diversi livelli(Fig. 1), cioè con una diminuzione della concentrazione di ormoni periferici (ovaie, in particolare estradiolo), la sintesi e il rilascio di ormoni dell'ipotalamo e della ghiandola pituitaria (rispettivamente ormone di rilascio delle gonadotropine (GnRH) e ormoni gonadotropici). Una caratteristica della regolazione della SM femminile è la presenza di un feedback positivo, quando in risposta ad un aumento significativo del livello di estradiolo nel follicolo preovulatoria aumenta la produzione di GnRH e gonadotropine (picco ovulatoria nel rilascio di LH e FSH ). Il funzionamento del sistema riproduttivo di una donna è caratterizzato da processi di regolazione ciclici (ripetitivi), le cui idee si adattano concetto moderno ciclo mestruale.
Il ciclo mestruale è un cambiamento ricorrente nell'attività del sistema ipotalamo-ipofisi-ovaio e nei cambiamenti strutturali e funzionali da essi causati. organi riproduttivi: utero, tube di Falloppio, ghiandole mammarie, vagina.
Il culmine di ogni ciclo è il sanguinamento mestruale (mestruazioni), il cui primo giorno è considerato l'inizio del ciclo mestruale. Il primo periodo nella vita di una ragazza si chiama menarca. età media menarca - 12-14 anni.
Riso. 1. Regolazione del sistema riproduttivo femminile: RG - ormoni di rilascio, FSH - ormone follicolo-stimolante, LH - ormone luteinizzante, TSH- ormone tireostimolante, ACTH - ormone adrenocorticotropo, Prl - prolattina, T4 - tiroxina, ADH - ormone antidiuretico, A - androgeni, E - estrogeni, P - progesterone, I - inibina, P - fattori di crescita; le frecce piene sono collegamenti diretti, le frecce tratteggiate sono collegamenti negativi inversi.
La durata del ciclo mestruale è determinata dal primo giorno di uno al primo giorno prossima mestruazione e normalmente varia da 21 a 35 giorni (negli adolescenti, entro 1,5-2 anni dal menarca, la durata del ciclo può essere più variabile - da 21 a 40-45 giorni). Un tale ciclo è chiamato normativo. Una variazione del ciclo normativo è ciclo ideale della durata di 28 giorni. Viene chiamato un ciclo mestruale ridotto (meno di 21 giorni). antiposizione (ciclo anteponente), allungamento (oltre 35 giorni) - postposizione (ciclo di post-posa).
La durata delle normali mestruazioni è in media di 3-5 giorni (normale - da 3 a 7 giorni) e la perdita media di sangue è di 50-70 ml (normale - fino a 80 ml).
Il ciclo mestruale è condizionatamente suddiviso in ovarico e cicli uterini. Ciclo ovarico (ovarico). implica processi ciclici che si verificano nelle ovaie sotto l'influenza di ormoni gonadotropici e di rilascio. I cambiamenti ciclici nel corpo di una donna sono carattere bifasico. Prima fase (follicolare, follicolare). il ciclo è determinato dalla maturazione del follicolo e dell'uovo nell'ovaio, dopodiché si rompe e l'uovo lo lascia - ovulazione. Seconda fase (luteale). associato alla formazione del corpo luteo. Simultaneamente in modo ciclico nell'endometrio si verificano in sequenza rigenerazione e proliferazione strato funzionale, mutevole attività secretoria le sue ghiandole terminano desquamazione strato funzionale (mestruazioni). I processi ciclici nell'endometrio sono fasi successive ciclo uterino.
Il significato biologico dei cambiamenti che si verificano durante il ciclo mestruale nelle ovaie e nell'endometrio è quello di garantire la funzione riproduttiva nelle fasi di maturazione dell'uovo, la sua fecondazione e l'impianto dell'embrione nell'utero. Se non si verifica la fecondazione dell'uovo, lo strato funzionale dell'endometrio viene respinto, compare una secrezione sanguinolenta dal tratto genitale e si verificano nuovamente e nella stessa sequenza nel sistema riproduttivo processi volti a garantire la maturazione dell'uovo.
Supremo V-esimo livello di regolamentazione ciclo mestruale è corteccia, vale a dire il sistema limbico e i nuclei dell'amigdaloide. La corteccia cerebrale esercita il controllo sul sistema ipotalamo-ipofisario attraverso i neurotrasmettitori (neurotrasmettitori), cioè trasmettitori impulso nervoso sui nuclei neurosecretori dell'ipotalamo. Il ruolo più importante è assegnato ai neuropeptidi (dopamina, norepinefrina, serotonina, kiss-peptina, la famiglia dei peptidi oppioidi), nonché all'ormone pineale melatonina. In situazioni stressanti, con un cambiamento del clima, del ritmo del lavoro (ad esempio i turni notturni), si osservano disturbi dell'ovulazione, che si realizzano attraverso un cambiamento nella sintesi e nel consumo di neurotrasmettitori nei neuroni cerebrali, nonché della melatonina nella ghiandola pineale.
Il SNC ha un gran numero di recettori per l'estradiolo e altri ormoni steroidei, che indica il loro ruolo importante non solo nell'implementazione del feedback, ma anche nel metabolismo dei neurotrasmettitori.
IVlivello del sistema riproduttivo - ipotalamo- è il più alto centro vegetativo, un ibrido del sistema nervoso ed endocrino, coordinando le funzioni di tutti organi interni e sistemi che mantengono l'omeostasi nel corpo. Sotto il controllo dell'ipotalamo c'è la ghiandola pituitaria e la regolazione ghiandole endocrine: gonadi (ovaie), ghiandola tiroidea, ghiandole surrenali (Fig. 1). Nell'ipotalamo ci sono due tipi di cellule neurosecretive che svolgono l'interazione ipotalamo-ipofisi:
Luogo di sintesi ormone di rilascio gonadotropico (GnRH) sono i nuclei arcuati dell'ipotalamo mediobasale. L'ormone di rilascio dell'LH, la luliberina, è stato isolato, sintetizzato e descritto. Ad oggi, non è stato possibile isolare e sintetizzare la folliberina. Pertanto, i liberini gonadotropici ipotalamici sono designati GnRH, poiché stimolano il rilascio sia di LH che di FSH dalla ghiandola pituitaria anteriore. La secrezione di GnRH è geneticamente programmata e avviene con un certo ritmo pulsante - 1 volta in 60-90 minuti (ritmo di secrezione circolare, oraria). Allo stato attuale, è stato dimostrato il ruolo permissivo (attivante) del GnRH nel funzionamento della SM. Il ritmo del polso della secrezione di GnRH si forma pubertà ed è un indicatore della maturità delle strutture neurosegrete dell'ipotalamo. Secrezione circolare di GnRH attiva il sistema ipotalamo-ipofisi-ovarico. Sotto l'influenza del GnRH, LH e FSH vengono rilasciati dalla ghiandola pituitaria anteriore.
La secrezione di GnRH è modulata dai neuropeptidi delle strutture extraipotalamiche, nonché dagli ormoni sessuali secondo il principio del feedback. In risposta ad un aumento del picco preovulatorio dell'estradiolo, aumenta la sintesi e il rilascio di GnRH, sotto l'influenza del quale aumenta la secrezione di gonadotropine, con conseguente ovulazione. Il progesterone ha sia un effetto inibitorio che stimolante sulla produzione di gonadotropine, agendo sul principio del feedback sia a livello dell'ipotalamo che a livello dell'ipofisi (Fig. 1).
Il ruolo principale nella regolazione del rilascio di prolattina appartiene alle strutture dopaminergiche dell'ipotalamo. La dopamina (DA) inibisce il rilascio di prolattina dalla ghiandola pituitaria, tireoliberina - stimola. Gli antagonisti della dopamina aumentano il rilascio di prolattina.
I neurosegreti dell'ipotalamo hanno un effetto biologico sul corpo in vari modi. Il percorso principale è paraipofisario attraverso le vene che scorrono nei seni della dura madre e da lì nella circolazione sistemica. Percorso transipofisario - attraverso il sistema della vena portale (portale) fino al lobo anteriore della ghiandola pituitaria; Una caratteristica del sistema circolatorio portale è la possibilità di flusso sanguigno al suo interno in entrambe le direzioni (sia all'ipotalamo che all'ipofisi), che è importante per l'implementazione dei meccanismi di feedback. L'effetto inverso sulla ghiandola pituitaria degli ormoni sessuali delle ovaie viene effettuato attraverso le arterie vertebrali.
Pertanto, la secrezione ciclica di GnRH innesca il sistema ipotalamo-ipofisi-ovarico, ma la sua funzione non può essere considerata autonoma; è modulata sia dai neuropeptidi del SNC che dagli steroidi ovarici in modo feedback.
IIIlivello - il lobo anteriore della ghiandola pituitaria (adenoipofisi). Nell'adenoipofisi si distinguono tre tipi di cellule: cromofobiche (riserva), acidofile e basofile. Qui vengono sintetizzati gli ormoni gonadotropici: ormone follicolo-stimolante o follitropina (FSH), luteinizzante o luteotropina (LH); così come la prolattina (Prl) e altri ormoni tropici: ormone stimolante la tiroide, tireotropina (TSH), ormone somatotropo (STH), ormone adrenocorticotropo, corticotropina (ACTH); melanostimolante, melanotropina (MSH) e ormone lipotropico (GPL). LH e FSH sono glicoproteine, Prl è un polipeptide.
La secrezione di LH e FSH è controllata(Fig. 1):
- GnRH, che entra nell'adenoipofisi attraverso il sistema portale e stimola la secrezione di gonadotropine;
- ormoni sessuali ovarici (estradiolo, progesterone) secondo il principio del feedback negativo o positivo;
- inibina A e B. L'inibina B è sintetizzata nelle ovaie e, insieme all'estradiolo, sopprime la secrezione di FSH nella seconda metà della fase follicolare del ciclo (dopo la selezione e la crescita del follicolo dominante). Con l'età, al diminuire del numero di follicoli, diminuisce la produzione di inibina B, che porta ad un progressivo aumento di FSH, che cerca di fornire livello normale estradiolo.
LH e FSH determinano i primi passi nella sintesi degli steroidi sessuali nelle ovaie interagendo con specifici recettori nei tessuti delle gonadi. L'efficacia della regolazione ormonale è determinata sia dalla quantità di ormone attivo che dal livello di contenuto del recettore nella cellula bersaglio.
Il ruolo biologico dell'FSH:
- crescita dei follicoli nelle ovaie, proliferazione delle cellule della granulosa nei follicoli;
- sintesi dell'aromatasi - enzimi che metabolizzano gli androgeni in estrogeni (produzione di estradiolo);
- sintesi dei recettori LH sulle cellule della granulosa del follicolo (preparazione per l'ovulazione);
- stimolazione della secrezione di attivina, inibina, fattori di crescita simili all'insulina (IGF), che svolgono un ruolo importante nella follicologenesi e nella sintesi degli steroidi sessuali.
Il ruolo biologico di LH:
- induce l'ovulazione (insieme all'FSH);
- sintesi di estradiolo nel follicolo dominante;
- sintesi degli androgeni nelle cellule della teca (cellule della guaina) del follicolo;
- luteinizzazione delle cellule della granulosa del follicolo ovulato e formazione di un corpo luteo;
- sintesi di progesterone e altri steroidi nelle cellule luteali del corpo luteo.
Prolattina (Prl)- un polipeptide sintetizzato dalle cellule dell'adenoipofisi (lattotrofi), controlla l'allattamento, stimola la crescita dei dotti mammari, supporta la funzione del corpo luteo e la sintesi del progesterone, ha vari effetti biologici: riduce la densità minerale ossea, aumenta l'attività delle cellule pancreatiche, porta all'insulino-resistenza (effetto diabetogeno), è coinvolta nella regolazione del metabolismo, del comportamento alimentare, dei cicli di sonno e veglia, della libido, ecc.
IIlivello del sistema riproduttivo - ovaie. La principale unità strutturale dell'ovaio è il follicolo contenente l'uovo (ovocita). Nelle ghiandole sessuali si verificano la crescita e la maturazione dei follicoli, l'ovulazione, la formazione del corpo luteo e la sintesi degli steroidi sessuali.
Processi follicologenesi nelle ovaie si verifica continuamente - dal periodo prenatale alla postmenopausa. Alla nascita, le ovaie di una ragazza contengono circa 2 milioni di follicoli primordiali (germinali primari). La maggior parte di loro subisce cambiamenti atretici (atresia - sviluppo inverso) per tutta la vita e solo una piccolissima parte attraversa un ciclo di sviluppo completo dal primordiale alla maturità con l'ovulazione e la successiva formazione del corpo luteo. Al momento del menarca, le ovaie contengono 200-450 mila follicoli primordiali (la cosiddetta riserva ovarica). Di questi, solo 400-500 possono ovulare durante la loro vita, il resto va incontro ad atresia (circa il 90%). Nel processo di atresia follicolare, un ruolo importante è svolto dall'apoptosi (morte cellulare programmata), un processo biologico che provoca il completo riassorbimento della cellula sotto l'influenza del proprio apparato lisosomiale. Durante un ciclo mestruale, di regola, si sviluppa solo un follicolo con un uovo all'interno. In caso di maturazione di un numero maggiore, è possibile la gravidanza multipla.
Un ruolo importante nei meccanismi di regolazione auto e paracrina della funzione non solo dell'ovaio, ma dell'intero sistema riproduttivo appartiene ai fattori di crescita.
Fattori di crescita (FR)- sostanze biologicamente attive che stimolano o inibiscono la differenziazione delle cellule che trasmettono un segnale ormonale. Sono sintetizzati in cellule aspecifiche di vari tessuti del corpo e hanno effetti autocrini, paracrini, intracrini ed endocrini. L'effetto autocrino si realizza influenzando le cellule sintetizzando direttamente questo FR. Paracrino - è realizzato dall'azione sulle cellule vicine. Effetto intracrino - RF agisce come un messaggero intracellulare (trasmettitore di segnale). L'effetto endocrino si realizza attraverso il flusso sanguigno alle cellule distanti.
Il ruolo più importante nella fisiologia del sistema riproduttivo è svolto dai seguenti RF: insulino-simile (IGF), epidermico (EGF), trasformante (TGF-α, TGF-β), fattore di crescita vascolare endoteliale (vasculoendoteliale) (VEGF ), inibine, attivine, ormone antimulleriano (AMG).
Fattori di crescita insulino-simili Ie II(IGF-I, IGF-II) sono sintetizzati nelle cellule della granulosa e in altri tessuti, stimolano la sintesi degli androgeni nelle cellule della teca ovarica, l'aromatizzazione degli androgeni in estrogeni, la proliferazione delle cellule della granulosa e la formazione dei recettori LH sulle cellule della granulosa. La loro produzione è regolata dall'insulina.
Fattore di crescita epidermico (EGF)- il più potente stimolatore della proliferazione cellulare, presente nelle cellule della granulosa, nello stroma endometriale, nelle ghiandole mammarie e in altri tessuti; ha un effetto oncogenico nei tessuti estrogeno-dipendenti (endometrio, ghiandole mammarie).
Fattore di crescita endoteliale vascolare (VEGF) svolge un ruolo importante nell'angiogenesi dei follicoli in crescita, così come nel mio e nell'endometrio. Il VEGF aumenta l'attività mitogenica delle cellule endoteliali, la permeabilità della parete vascolare. L'espressione del VEFR è aumentata nell'endometriosi, nel mioma uterino, nei tumori delle ovaie e delle ghiandole mammarie, nella PCOS, ecc.
Fattori di crescita trasformanti (TGF-α, TGF-β) stimolare la proliferazione cellulare, partecipare alla crescita e alla maturazione dei follicoli, alla proliferazione delle cellule della granulosa; hanno un effetto mitogenico e oncogenico, la loro espressione è aumentata nel cancro dell'endometrio e dell'ovaio. Le sostanze proteiche della famiglia del TGF-β includono le inibine, l'attivina, la follistatina e l'AMH.
Inibizioni (A e B)- le sostanze proteiche, formate nelle cellule della granulosa e in altri tessuti, sono coinvolte nella regolazione della sintesi di FSH, inibendola, come l'estradiolo, con un meccanismo di feedback simile. La formazione di inibina B aumenta a metà della fase follicolare del ciclo parallelamente all'aumento delle concentrazioni di estradiolo dopo la selezione del follicolo dominante e, raggiungendo un massimo, inibisce il rilascio di FSH.
Attivo presente nelle cellule della granulosa del follicolo e dei gonadotropi ipofisari, stimola la sintesi dell'FSH, la proliferazione delle cellule della granulosa, l'aromatizzazione degli androgeni in estrogeni, inibisce la sintesi degli androgeni nelle cellule della teca, previene la luteinizzazione spontanea (prematura, prima dell'ovulazione) del follicolo preovulatorio , stimola la produzione di progesterone nel corpo luteo.
Follistatina- proteina bloccante l'FSH, secreta dalle cellule della ghiandola pituitaria anteriore, la granulosa; sopprime la secrezione di FSH.
Ormone antimulleriano (AMH)- un rappresentante della famiglia TGF-β, prodotto nelle donne nelle cellule della granulosa dei follicoli preantrali e piccoli antrali, svolge un ruolo importante nei meccanismi di reclutamento e selezione dei follicoli, è un indicatore quantitativo della riserva ovarica e viene utilizzato nella pratica clinica per valutarlo e predire la risposta ovarica alla stimolazione dell'ovulazione e può anche fungere da marker di tumori a cellule della granulosa delle ovaie, in cui l'AMH è significativamente aumentato. L'AMH non è controllato dalle gonadotropine, non è coinvolto nel classico ciclo di feedback (a differenza di FSH, estradiolo e inibina B), non dipende dalla fase del ciclo e agisce come fattore paracrino nella regolazione del sistema riproduttivo.
Follicogenesi nelle ovaie
Nell'ovaio di una donna, i follicoli sono a vari stadi di maturità. La follicologenesi inizia dalla 12a settimana di sviluppo prenatale; la maggior parte dei follicoli subisce atresia. Non è del tutto noto quali fattori siano responsabili della crescita dei follicoli primordiali. Follicoli primordiali caratterizzato da un unico strato di cellule pregranulari piatte, un piccolo ovocita immaturo (che non ha completato la seconda divisione della meiosi), le cellule della teca (conchiglie) sono assenti.
Fasi di crescita del follicolo:
- Prima fase di crescita – follicoli primordiali a preantrali– crescita non ormonale dipendente(non dipende da FSH). Dura circa 3-4 mesi, fino alla formazione di follicoli con un diametro di 1-4 mm. A follicoli preantrali primari c'è uno strato di cellule della granulosa, l'ovocita inizia ad aumentare, appare la teca. Follicoli preantrali secondari caratterizzato da 2-8 strati
- Seconda fase - crescita dei follicoli preantrali allo stadio dei follicoli antrali. Richiede circa 70 giorni e si verifica in presenza di concentrazioni minime di FSH - stadio ormono-dipendente della crescita del follicolo. Anche IPFR-I e AMH svolgono un ruolo importante in questa fase. Follicoli antrali hanno una cavità piena di liquido al centro, il loro diametro all'inizio del ciclo mestruale è di 3-4 mm (determinato dall'ecografia in qualsiasi giorno del ciclo mestruale), tendono a crescere rapidamente nella fase follicolare precoce (Fig. 2, 3).

Riso. 2. Fasi dello sviluppo del follicolo

Pertanto, la durata totale della follicologenesi dal momento dell'inizio della crescita dei follicoli primordiali all'ovulazione di un follicolo maturo è di circa 200 giorni; sul fase follicolare il successivo ciclo mestruale rappresenta solo la fase finale della formazione del follicolo dominante e dell'ovulazione. Poiché i processi di follicologenesi si verificano continuamente, questo può spiegare la presenza di follicoli nelle ovaie. varie fasi maturità, determinata ecograficamente, in qualsiasi giorno del ciclo mestruale (Fig. 3).
ciclo ovarico si compone di due fasi: follicolare e luteale. Conto alla rovescia fase follicolare il ciclo inizia il primo giorno della mestruazione successiva, con un ciclo mestruale ideale, la prima fase dura circa 2 settimane, è caratterizzata dalla crescita e maturazione del follicolo dominante e termina con la sua ovulazione, che avviene il 13-14° giorno del ciclo Poi viene fase luteale ciclo che dura da 14-15 a 28 giorni, durante il quale avviene la formazione, lo sviluppo e la regressione del corpo luteo. In un ciclo anteponico o posticipato, la durata della fase follicolare può differire da quella di un ciclo ideale o prossimo all'ideale.
Fase follicolare del ciclo ovarico.
La crescita del follicolo gonadotropina-dipendente inizia alla fine del ciclo mestruale precedente. Secondo il principio, si verifica un aumento della sintesi e del rilascio di FSH da parte della ghiandola pituitaria feedback negativo in risposta a una diminuzione del livello di progesterone, estradiolo e inibina B con regressione del corpo luteo. Sotto l'influenza dell'FSH, la crescita dei follicoli antrali continua e nella prima fase follicolare del ciclo mestruale (4-5 giorni dall'inizio delle mestruazioni), le loro dimensioni sono di 4-5 mm di diametro. Durante questo periodo, l'FSH stimola la proliferazione e la differenziazione delle cellule della granulosa, la sintesi dei recettori LH in esse, l'attivazione dell'aromatasi e la sintesi degli estrogeni e dell'inibina. L'LH nella fase follicolare precoce colpisce principalmente la sintesi degli androgeni - precursori degli estrogeni.
L'FSH raggiunge il suo valore massimo entro il 5-6° giorno del ciclo mestruale, dopodiché diminuisce (sotto l'influenza di concentrazioni crescenti di estradiolo e inibina B, sintetizzate dalla granulosa dei follicoli antrali in crescita), quindi aumenta di nuovo contemporaneamente con LH a il picco ovulatoria nel ciclo 13-14 giorni (Fig. 4). Selezione del follicolo dominante avviene entro il 5-7° giorno del ciclo da un pool di follicoli antrali con un diametro di 5-10 mm. Dominante diventa il follicolo con il diametro maggiore, con numero più grande cellule della granulosa e recettori dell'FSH, grazie ai quali il follicolo dominante mantiene la capacità di crescere ulteriormente e sintetizzare l'estradiolo nonostante una diminuzione del livello di FSH nel sangue. L'ulteriore crescita del follicolo dominante, a partire dalla metà della fase follicolare del ciclo, diventa non solo FSH-dipendente, ma anche LH- e FSH-dipendente. Nella rapida crescita del follicolo principale, giocano un ruolo anche le crescenti concentrazioni di estradiolo e FR - IGF, SEFR. Al momento dell'ovulazione, il follicolo dominante raggiunge una dimensione di 18-21 mm (Fig. 3). Nei restanti follicoli antrali, una diminuzione del livello sierico di FSH provoca processi di atresia (apoptosi). Nei meccanismi di atresia dei follicoli immaturi, un certo ruolo è assegnato ad alte concentrazioni di androgeni sintetizzati negli stessi piccoli follicoli (Fig. 2, 3).
Ovulazione- rottura di un follicolo maturo e rilascio di un uovo da esso. Il processo di ovulazione si verifica quando livello massimo di estradiolo nel follicolo preovulatoria (Fig. 4), che, secondo riscontro positivo stimola il rilascio ovulatoria di LH e FSH da parte della ghiandola pituitaria. L'ovulazione avviene 10-12 ore dopo il picco di LH o 24-36 ore dopo il picco di estradiolo (Fig. 4). Il processo di rottura della membrana basale del follicolo avviene sotto l'influenza di vari enzimi e sostanze biologicamente attive nelle cellule della granulosa luteinizzata: enzimi proteolitici, plasmina, istamina, collagenasi, prostaglandine, ossitocina e relaxina. L'importante ruolo del progesterone, che è sintetizzato nelle cellule luteinizzate del follicolo preovulatorio sotto l'influenza del picco di LH, ha dimostrato di svolgere un ruolo importante nell'attivazione degli enzimi proteolitici coinvolti nella rottura della membrana basale del follicolo . L'ovulazione è accompagnata da sanguinamento dai capillari rotti che circondano le cellule della teca.

Fase luteale del ciclo ovarico
Dopo l'ovulazione, i capillari formati crescono rapidamente nella cavità del follicolo ovulato, le cellule della granulosa subiscono ulteriormente luteinizzazione con la formazione di un corpo luteo che secerne progesterone sotto l'influenza di LH. La luteinizzazione delle cellule della granulosa si manifesta morfologicamente in un aumento del loro volume e nella formazione di inclusioni lipidiche. corpo luteo - formazione transitoria ormonale attiva, funzionante per 14 giorni, indipendentemente dalla durata totale del ciclo mestruale. Un vero e proprio corpo luteo si sviluppa solo nella fase in cui nel follicolo preovulatore si forma un numero adeguato di cellule della granulosa con un alto contenuto di recettori LH. Nello sviluppo del corpo luteo si distinguono i seguenti fasi:
- proliferazione- caratterizzato da luteinizzazione attiva delle cellule della granulosa sotto l'influenza di LH;
- vascolarizzazione- germinazione dei capillari nel corpo luteo;
- periodo d'oro- questa fase cade nei giorni 21-22 del ciclo, caratterizza il completamento della formazione strutturale del corpo luteo, a cui corrisponde un progressivo aumento delle concentrazioni di steroidi sessuali (Fig. 4); l'azione combinata di progesterone ed estradiolo contribuisce alla preparazione preimpianto dell'endometrio (trasformazione secretoria);
- sviluppo inverso (regressione)- diminuzione dell'attività del corpo luteo, associata ad una diminuzione del numero di recettori per l'LH; un effetto luteolitico è esercitato anche da elevate concentrazioni di estradiolo e Prl al termine del ciclo mestruale; la regressione del corpo luteo porta ad una diminuzione del livello di progesterone (Fig. 4), che provoca la desquamazione dell'endometrio nell'utero - il ciclo si ripete.
Se si verifica il concepimento e l'impianto dell'ovulo (nei giorni 21-22 del ciclo), il corion emergente inizia a produrre gonatropina corionica umana (hCG), che stimola l'ulteriore sviluppo del corpo luteo. In questo caso, è formato corpo giallo della gravidanza che continua a sintetizzare il progesterone in alte concentrazioni necessarie per prolungare la gravidanza. Il corpo luteo della gravidanza esiste fino a 8-10 settimane di gestazione, quindi subisce una regressione e supporto ormonale la gravidanza prende il sopravvento sulla placenta formata alla fine del 1o trimestre.
Funzione ormonale delle ovaie
I processi ciclici nell'ovaio sono caratterizzati non solo da cambiamenti morfologici nei follicoli e nel corpo luteo, ma anche dai processi di steroidogenesi, la formazione di ormoni sessuali, che sono indissolubilmente legati ad essi. Attualmente è generalmente accettato teoria delle due cellule biosintesi degli steroidi nelle ovaie, secondo cui l'LH stimola la sintesi degli androgeni nelle cellule della teca, mentre l'FSH stimola la sintesi degli enzimi aromatasi che metabolizzano gli androgeni in estrogeni nelle cellule della granulosa.
Le strutture delle ovaie che producono steroidi sono la granulosa, la teca e, in misura minore, le cellule dello stroma. Le cellule della teca sono la principale fonte di androgeni, cellule della granulosa - estrogeni, il progesterone è sintetizzato nelle cellule della teca e al massimo nelle cellule luteali del corpo luteo (cellule della granulosa luteinizzata). Il substrato per tutti gli steroidi, compresi quelli surrenali e testicolari, è il colesterolo (Fig. 5).

La sintesi degli ormoni sessuali avviene anche extragonadalmente. È noto che nel tessuto adiposo è presente un sistema enzimatico P450 aromatasi, che è coinvolto nella conversione degli androgeni in estrogeni. Questo processo può essere avviato da vari RF mitogeni o dall'estradiolo stesso. Inoltre, il testosterone biologicamente attivo (diidrotestosterone) viene anche sintetizzato extragonadalmente nei tessuti bersaglio periferici ( follicoli piliferi, ghiandole sebacee) sotto l'influenza dell'enzima 5-α-reduttasi.
Circa il 96% di tutti gli steroidi sessuali sono in uno stato legato alle proteine, in particolare, con globulina legante gli steroidi sessuali (SHBG) così come le albumine, la cui sintesi viene effettuata nel fegato. L'effetto biologico degli ormoni è determinato da frazioni libere non legate, il cui livello cambia in varie condizioni patologiche, in particolare insulino-resistenza, malattie del fegato, ecc.
Estrogeni. Le principali frazioni di estrogeni sono estrone (E 1 ), estradiolo (E 2 ), estriolo (E 3 ). Il più biologicamente attivo è l'estradiolo. L'estriolo è un metabolita periferico dell'estrone e dell'estradiolo e non un prodotto indipendente della secrezione ovarica. Nel 1965 fu descritto anche un quarto estrogeno: estetrolo (E 4 ), finora poco studiato, con un debole effetto estrogenico.
Azione biologica degli estrogeni:
- sul riproduttivo organi bersaglio:
- proliferazione di endo e miometrio, epitelio vaginale, cervice;
- secrezione di muco nel canale cervicale;
- crescita dei dotti delle ghiandole mammarie;
- sul non riproduttivo tessuti bersaglio:
- processi proliferativi della mucosa dell'uretra, vescica;
- sviluppo del sistema muscolo-scheletrico, aumento della mineralizzazione ossea (dovuta alla stimolazione della sintesi degli osteoblasti);
- diminuzione della secrezione delle ghiandole sebacee;
- aumento della sintesi e della maturazione del collagene nella pelle;
- riduzione dell'irsutismo (effetto antiandrogeno dovuto a una diminuzione della clearance di SHPS);
- effetto anti-aterogeno (riduzione delle frazioni lipidiche aterogeniche);
- la distribuzione del tessuto adiposo e la formazione dello scheletro secondo il tipo femminile, il timbro della voce femminile;
- miglioramento delle funzioni del sistema nervoso centrale (cognitivo, ecc.);
- effetto protettivo sull'endotelio vascolare (effetto antiaterosclerotico);
- aumento delle proprietà di coagulazione del sangue, trombosi (a causa dell'aumentata sintesi dei fattori della coagulazione nel fegato);
- aumento della libido.
L'effetto biologico degli estrogeni su vari organi e tessuti dipende dal numero e dal tipo di recettori specifici e dalla loro sensibilità. È stata accertata la presenza di due tipi di recettori per l'estradiolo: ER- α - recettori nucleari avere un effetto proliferativo, e membrana ER- β , avere un effetto antiproliferativo.
Gestageni. Il principale gestageno è il progesterone, che si forma principalmente nel corpo luteo delle ovaie.
L'azione biologica del progesterone:
L'azione del progesterone si realizza attraverso i recettori tipo A e B. A seconda della prevalenza dell'uno o dell'altro tipo di recettore, i tessuti bersaglio rispondono con effetti diversi. Ad esempio, nell'endometrio e nell'epitelio delle ghiandole mammarie, PR tipo A , quindi il progesterone se ne rende conto azione antiproliferativa(gli analoghi del progesterone sono ampiamente utilizzati per il trattamento e la prevenzione dei processi iperplastici dell'endometrio e delle ghiandole mammarie, mastopatia fibrocistica). Il miometrio è dominato da PR tipo B e spettacoli di progesterone effetto proliferativo. Pertanto, secondo i concetti moderni, svolge un ruolo importante nella patogenesi dei fibromi uterini e nel trattamento di questo tumore vengono utilizzati con successo modulatori PR selettivi che bloccano i recettori di tipo B.Androgeni. Le principali frazioni di androgeni sono androgeni forti testosterone, il suo debole predecessore androstenedione, così come diidroandrostenedione (DHEA) e suo solfato (DHEA-S). Il metabolita biologicamente più attivo del testosterone è diidrotestosterone, sintetizzato nei tessuti bersaglio periferici (follicoli piliferi, ghiandole sebacee) sotto l'influenza dell'enzima 5-α-reduttasi. I principali siti di sintesi degli androgeni nel corpo femminile sono le ovaie, le ghiandole surrenali, nonché il tessuto adiposo e la pelle con le sue appendici.
Effetti biologici degli androgeni:
iolivello regolazione della funzione riproduttiva sono sensibili alle fluttuazioni dei livelli di steroidi sessuali nelle parti interne ed esterne del sistema riproduttivo (utero, le tube di Falloppio, mucosa vaginale), nonché ghiandole mammarie. I cambiamenti ciclici più pronunciati si verificano nell'endometrio e costituiscono il ciclo uterino.ciclo uterino
I cambiamenti ciclici nell'endometrio lo influenzano strato superficiale funzionale, costituito da cellule epiteliali compatte e intermedie, che vengono respinte durante le mestruazioni. strato basale, non respinto durante le mestruazioni, assicura il ripristino degli strati desquamati.
Le trasformazioni cicliche dello strato funzionale dell'endometrio procedono secondo il ciclo ovarico in tre fasi successive. – fase di proliferazione, fase di secrezione e fase di desquamazione (mestruazioni).
fase di desquamazione. Il sanguinamento mestruale osservato alla fine di ogni ciclo mestruale è dovuto al rigetto dello strato funzionale dell'endometrio. L'inizio delle mestruazioni è considerato il primo giorno del ciclo mestruale. La durata del sanguinamento mestruale è in media di 3-5 giorni. A causa della regressione del corpo luteo e di una forte diminuzione del contenuto di steroidi sessuali nell'endometrio, l'ipossia aumenta. L'inizio delle mestruazioni è facilitato da uno spasmo prolungato delle arterie, che porta alla stasi del sangue e alla formazione di coaguli di sangue. ipossia tissutale ( acidosi tissutale) esacerbare l'aumentata permeabilità dell'endotelio, la fragilità delle pareti dei vasi sanguigni, numerosi piccole emorragie e massiccia infiltrazione di leucociti. Gli enzimi proteolitici lisosomiali rilasciati dai leucociti favoriscono la fusione degli elementi tissutali. A seguito di uno spasmo prolungato dei vasi, la loro espansione paretica si verifica con un aumento del flusso sanguigno. Allo stesso tempo, si nota un aumento della pressione idrostatica nel microcircolo e una rottura delle pareti dei vasi, che a questo punto hanno in gran parte perso la loro resistenza meccanica. In questo contesto, si verifica la desquamazione attiva delle aree necrotiche dello strato funzionale. Entro la fine del 1° giorno delle mestruazioni, 2/3 dello strato funzionale viene respinto e la sua completa desquamazione di solito termina il 3° giorno.
Il flusso mestruale contiene sangue e muco cervicale, ricco di leucociti. Il sangue mestruale difficilmente coagula, è ricco di ioni calcio, contiene poco fibrinogeno e manca di protrombina. In media, una donna perde 50-70 ml di sangue durante le mestruazioni.
Immediatamente dopo il rigetto dell'endometrio necrotico, fase di rigenerazione , caratterizzato dall'epitelizzazione della superficie della ferita dell'endometrio a causa delle cellule dello strato basale. I processi di rigenerazione avvengono sotto il controllo degli estrogeni e contribuiscono, insieme al vasospasmo e alla formazione di trombi, a fermare il sanguinamento mestruale. Alcuni autori individuano la rigenerazione come una fase separata del ciclo uterino.
fase di proliferazione. Desquamazione e rigenerazione della mucosa al termine delle mestruazioni entro il 3°-5° giorno del ciclo. Quindi, sotto l'influenza di una crescente concentrazione di estrogeni, lo spessore dello strato funzionale aumenta a causa della crescita di tutti gli elementi dello strato basale: ghiandole, stroma, vasi sanguigni. Le ghiandole endometriali hanno la forma di tubuli diritti o più contorti con un lume diretto. Le arterie spirali sono leggermente tortuose. In scena proliferazione tardiva(11-14 giorni del ciclo), le ghiandole endometriali diventano contorte, arrotondate a forma di cavatappi, il loro lume è leggermente espanso. Le arterie a spirale che crescono dallo strato basale raggiungono la superficie dell'endometrio, sono alquanto tortuose. Lo spessore dello strato funzionale dell'endometrio entro la fine della fase di proliferazione raggiunge 7-8 mm.
Fase di secrezione (trasformazione secretoria) inizia dopo l'ovulazione il 13-14° giorno del ciclo, dura 14 giorni ed è direttamente correlato all'attività del corpo luteo. È caratterizzato dal fatto che l'epitelio delle ghiandole sotto l'influenza del progesterone e dell'estradiolo inizia a produrre un segreto contenente glicosaminoglicani acidi, glicoproteine, glicogeno.
A fase iniziale fasi di secrezione (15-18° giorno del ciclo) compaiono i primi segni di trasformazioni secretorie. Le ghiandole diventano più tortuose, il loro lume è leggermente espanso. Negli strati superficiali dell'endometrio possono esserci emorragie focali associate a una diminuzione a breve termine degli estrogeni dopo l'ovulazione.
A fase intermedia fasi di secrezione (19-23 giorni di ciclo), quando la concentrazione di progesterone è massima e il livello di estrogeni aumenta, lo strato funzionale dell'endometrio diventa più alto (9-12 mm) e si divide nettamente in 2 strati. Strato profondo (spugnoso, spugnoso), confinante con la base, contiene un gran numero di ghiandole altamente contorte e una piccola quantità di stroma. Strato denso (compatto). è - 1/4-1/5 dello spessore dello strato funzionale. Ha meno ghiandole e più cellule del tessuto connettivo. La secrezione è più pronunciata nei giorni 20-21 del ciclo. A questo punto, nello stroma dell'endometrio si verificano trasformazioni simili a decidua (le cellule dello strato compatto diventano grandi, di forma arrotondata o poligonale, il glicogeno appare nel loro citoplasma). Le arterie a spirale sono fortemente tortuose, formano "grovigli" e si trovano nell'intero strato funzionale, la permeabilità vascolare aumenta, i lumi vascolari si espandono e il volume di afflusso di sangue all'endometrio aumenta. Questi cambiamenti nelle ghiandole e nei vasi dell'endometrio sono l'essenza della sua preparazione pre-impianto e sono sincronizzati nel tempo con l'ingresso dell'uovo fetale nella cavità uterina (la cosiddetta finestra di impianto è il 7° giorno dopo il concepimento). Se l'impianto ha successo, l'endometrio subirà una trasformazione decidua sotto l'influenza di una concentrazione crescente di progesterone. In assenza di gravidanza, si verificano alterazioni degenerative dell'endometrio.
Fase avanzata della fase di secrezione (24-27 giorni del ciclo) caratterizzato da una violazione del trofismo dell'endometrio e da un graduale aumento dei suoi cambiamenti degenerativi. L'altezza dell'endometrio diminuisce, lo stroma dello strato funzionale si restringe, la piegatura delle pareti delle ghiandole aumenta e acquisiscono contorni stellati o a dente di sega. Il 26-27° giorno del ciclo si osservano dilatazioni lacunari dei capillari ed emorragie focali nello stroma negli strati superficiali dello strato compatto. Viene chiamato lo stato dell'endometrio, così preparato alla disintegrazione e al rigetto mestruazioni anatomiche e viene rilevato un giorno prima dell'inizio mestruazioni cliniche(sanguinamento).
membrana mucosa istmo dell'utero nella struttura morfologica è simile all'endometrio, tuttavia non distingue tra lo strato funzionale e quello basale.
Nel canale cervicale si verificano anche cambiamenti ciclici. Durante le mestruazioni, la desquamazione non si verifica sulla membrana mucosa del canale cervicale, ma solo sul suo epitelio superficiale. Sotto l'influenza degli estrogeni nella fase follicolare del ciclo, il canale cervicale si espande, il sistema operativo esterno si apre leggermente (positivo "sintomo pupillare"), la produzione di muco cervicale aumenta, raggiungendo il massimo al momento dell'ovulazione (positivo "sintomo felce" , "sintomo di tensione del muco cervicale" - 8-10 cm). Sotto l'influenza del progesterone nella fase luteale del ciclo, il canale cervicale si restringe, la faringe esterna si chiude (negativo
"Sintomo della pupilla"), il muco cervicale diventa denso, denso, non si allunga (Tabella 1), la mucosa della cervice, la vagina diventa cianotica.
Si verificano cambiamenti ciclici mucosa della vagina, che è rappresentato da epitelio squamoso stratificato non cheratinizzato, quindi nella prima metà del ciclo, sotto l'influenza degli estrogeni
vi è una proliferazione degli strati intermedi e superficiali della mucosa. Nello striscio vaginale predominano le cellule mature e superficiali, l'indice karyopyknotic (KPI) è alto - 60-80% nel periodo preovulatoria (Tabella 1). Nella seconda fase del ciclo, sotto l'influenza del progesterone, si verifica l'apoptosi e la desquamazione delle cellule di superficie. Le cellule intermedie predominano nello striscio, assumono una forma allungata e si trovano principalmente in gruppi (indice di affollamento; CPI è basso - 20-25%, vedere Tabella 1).
Tabella 1. Prove diagnostica funzionale

Nota: TFD - test di diagnostica funzionale, KPI - indice cariopiconotico, BT - temperatura corporea basale; giorni del ciclo mestruale: 0 - giorno dell'ovulazione, numeri con segno "-" - giorni prima dell'ovulazione (fase follicolare del ciclo), numeri con segno "+" - giorni dopo l'ovulazione (fase luteale del ciclo).
Nelle ghiandole mammarie sotto l'influenza degli estrogeni nella prima metà del ciclo mestruale si verifica la proliferazione dell'epitelio dei passaggi lattiferi e nella seconda fase, sotto l'influenza del progesterone, la proliferazione dell'epitelio secretorio negli acini (lobuli).
Visualizza e acquista libri sull'ecografia Medvedev:
Ciclo mestrualeè un complesso di complessi processi biologici che si verificano nel corpo di una donna, caratterizzato da cambiamenti ciclici in tutte le parti del sistema riproduttivo ed è progettato per garantire il concepimento e lo sviluppo della gravidanza.
Le mestruazioni sono un breve sanguinamento uterino ciclico risultante dal rigetto dello strato funzionale dell'endometrio alla fine di un ciclo mestruale in due fasi. Il primo giorno delle mestruazioni è considerato il primo giorno del ciclo mestruale.
La durata del ciclo mestruale è il tempo che intercorre tra i primi giorni delle ultime due mestruazioni e varia normalmente da 21 a 36 giorni, in media - 28 giorni; durata delle mestruazioni - da 2 a 7 giorni; il volume della perdita di sangue è di 40-150 ml.
Fisiologia del sistema riproduttivo femminile
La regolazione neuroumorale del sistema riproduttivo è organizzata secondo un principio gerarchico. Si distingue
cinque livelli, ciascuno dei quali è regolato dalle strutture sovrastanti dal meccanismo di feedback: corteccia cerebrale, ipotalamo, ghiandola pituitaria, ovaie, utero e altri tessuti bersaglio per gli ormoni sessuali.
Corteccia
Il più alto livello di regolazione è la corteccia cerebrale: neuroni specializzati ricevono informazioni sullo stato dell'ambiente interno ed esterno, le convertono in segnali neuroumorali, che attraverso il sistema dei neurotrasmettitori entrano nelle cellule neurosensoriali dell'ipotalamo. La funzione dei neurotrasmettitori è svolta da ammine-catecolamine biogene - dopamina e norepinefrina, indoli - serotonina e neuropeptidi oppioidi - endorfine ed encefaline.
La dopamina, la noradrenalina e la serotonina esercitano il controllo sui neuroni ipotalamici che secernono l'ormone di rilascio delle gonadotropine (GnRH): la dopamina supporta la secrezione di GnRH nei nuclei arcuati e inibisce anche il rilascio di prolattina da parte dell'adenoipofisi; la norepinefrina regola la trasmissione degli impulsi ai nuclei prebiotici dell'ipotalamo e stimola il rilascio ovulatoria di GnRH; la serotonina controlla la secrezione ciclica dell'ormone luteinizzante (LH). I peptidi oppioidi sopprimono la secrezione di LH, inibiscono l'effetto stimolante della dopamina e il loro antagonista, il naloxone, provoca un forte aumento dei livelli di GnRH.
Ipotalamo
L'ipotalamo è una delle principali formazioni del cervello coinvolte nella regolazione del sistema nervoso autonomo, viscerale, trofico e neuro funzioni endocrine. I nuclei della zona ipofisi-tropica dell'ipotalamo (sopraottica, paraventricolare, arcuata e ventromediale) producono specifici neurosegreti con effetto farmacologico diametralmente opposto: rilascio di ormoni che liberano ormoni tropici nell'ipofisi anteriore e statine che ne inibiscono il rilascio.
Attualmente sono noti 6 ormoni di rilascio (RG): RG gonadotropico, RG stimolante la tiroide, RG adrenocorticotropo, RG somatotropico, RG melanotropico, prolattina-RG e tre statine: ormone inibitorio melanotropico, somatotropina
Ormone inibitorio di Ropny, ormone inibente la prolattina.
Il GnRH viene rilasciato nella circolazione portale in una modalità pulsante: 1 volta in 60-90 minuti. Questo ritmo è chiamato circo-rale. La frequenza del rilascio di GnRH è programmata geneticamente. Durante il ciclo mestruale, cambia entro piccoli limiti: la frequenza massima viene registrata nel periodo preovulatorio, il minimo - nella II fase del ciclo.
pituitario
Le cellule basofile dell'adenoipofisi (gonadotropociti) secernono ormoni - gonadotropine, che sono direttamente coinvolte nella regolazione del ciclo mestruale; questi includono: follitropina o ormone follicolo-stimolante (FSH) e lutropina o ormone luteinizzante (LH); un gruppo di cellule acidofile dell'ipofisi anteriore - i lattotropociti producono prolattina (PRL).
La secrezione di prolattina segue un ritmo di rilascio circadiano.
Esistono due tipi di secrezione di gonadotropine: tonica e ciclica. Il rilascio tonico di gonadotropine favorisce lo sviluppo dei follicoli e la loro produzione di estrogeni; ciclico - prevede un cambiamento nelle fasi di bassa e alta secrezione di ormoni e, in particolare, il loro picco preovulatoria.
Azione biologica dell'FSH: stimola la crescita e la maturazione dei follicoli, la proliferazione delle cellule della granulosa; induce la formazione di recettori LH sulla superficie delle cellule della granulosa; aumenta il livello di aromatasi nel follicolo in maturazione.
Azione biologica dell'LH: stimola la sintesi degli androgeni (precursori degli estrogeni) nelle cellule della teca; attiva l'azione delle prostaglandine e degli enzimi proteolitici, che portano all'assottigliamento e alla rottura del follicolo; si verifica la luteinizzazione delle cellule della granulosa (formazione del corpo luteo); Insieme al PRL, stimola la sintesi del progesterone nelle cellule della granulosa luteinizzata del follicolo ovulato.
L'effetto biologico del PRL: stimola la crescita delle ghiandole mammarie e regola l'allattamento; ha un effetto mobilizzante e ipotensivo; in quantità maggiori inibisce la crescita e la maturazione del follicolo; partecipa alla regolazione della funzione endocrina del corpo luteo.
ovaie
La funzione generativa delle ovaie è caratterizzata dalla maturazione ciclica del follicolo, dall'ovulazione, dal rilascio di un ovulo in grado di concepire e dalla fornitura delle trasformazioni secretorie nell'endometrio necessarie per la percezione di un ovulo fecondato.
La principale unità morfofunzionale delle ovaie è il follicolo. Secondo l'International Histological Classification (1994), si distinguono 4 tipi di follicoli: primordiale, primario, secondario (antrale, cavitario, vescicolare), maturo (preovulatoria, Graaf).
I follicoli primordiali si formano nel quinto mese di sviluppo fetale (a causa della meiosi, contengono un insieme aploide di cromosomi) e continuano ad esistere per tutta la vita di una donna fino alla menopausa e per diversi anni dopo la cessazione persistente delle mestruazioni. Al momento della nascita, entrambe le ovaie contengono circa 300-500 mila follicoli primordiali, in seguito il loro numero diminuisce drasticamente e all'età di 40 anni è di circa 40-50 mila a causa dell'atresia fisiologica.
Il follicolo primordiale è costituito da un ovulo circondato da un'unica fila di epitelio follicolare; il suo diametro non supera i 50 micron.
Lo stadio del follicolo primario è caratterizzato da una maggiore riproduzione dell'epitelio follicolare, le cui cellule acquisiscono una struttura granulare e formano un granulare (strato granulare). Il segreto secreto dalle cellule di questo strato si accumula nello spazio intercellulare. La dimensione dell'uovo aumenta gradualmente fino a 55-90 micron di diametro.
Nel processo di formazione del follicolo secondario, le sue pareti vengono allungate dal liquido: l'ovocita in questo follicolo non aumenta più (a momento presente il suo diametro è di 100-180 micron), tuttavia il diametro del follicolo stesso aumenta ed è di 20-24 mm.
In un follicolo maturo, l'ovulo, racchiuso nell'ovidotto, è ricoperto di guscio trasparente, su cui le cellule granulari si trovano nella direzione radiale e formano una corona radiante.
Ovulazione - rottura di un follicolo maturo con il rilascio di un ovulo circondato da una corona radiosa nella cavità addominale,
e più avanti nell'ampolla della tuba di Falloppio. La violazione dell'integrità del follicolo si verifica nella parte più convessa e sottile di esso, chiamata stigma.
In una donna sana, un follicolo matura durante il ciclo mestruale e per l'intero periodo riproduttivo ovula circa 400 uova, i restanti ovociti subiscono atresia. La vitalità dell'uovo viene mantenuta per 12-24 ore.
La luteinizzazione è una specifica trasformazione del follicolo nel periodo postovulatoria. Come risultato della luteinizzazione (colorazione gialla a causa dell'accumulo di pigmento lipocromico - luteina), della riproduzione e della crescita delle cellule della membrana granulare del follicolo ovulato, si forma una formazione chiamata corpo luteo. Nei casi in cui non avviene la fecondazione, il corpo luteo esiste per 12-14 giorni e poi subisce uno sviluppo inverso.
Pertanto, il ciclo ovarico consiste in due fasi: follicolina e luteale. La fase follicolina inizia dopo le mestruazioni e termina con l'ovulazione; la fase luteale occupa l'intervallo tra l'ovulazione e l'inizio delle mestruazioni.
Funzione ormonale delle ovaie
Le cellule della membrana della granulosa, il rivestimento interno del follicolo e il corpo luteo durante la loro esistenza svolgono la funzione di una ghiandola endocrina e sintetizzano tre tipi principali di ormoni steroidei: estrogeni, gestageni e androgeni.
Gli estrogeni sono secreti dalle cellule della membrana granulare, dal rivestimento interno e, in misura minore, dalle cellule interstiziali. A piccola quantità gli estrogeni si formano nel corpo luteo, lo strato corticale delle ghiandole surrenali, nelle donne in gravidanza - nella placenta. I principali estrogeni dell'ovaio sono estradiolo, estrone ed estriolo (i primi due ormoni sono prevalentemente sintetizzati). L'attività di 0,1 mg di estrone è assunta come 1 UI di attività estrogenica. Secondo il test di Allen e Doisy (la quantità più piccola del farmaco che causa l'estro nei topi castrati), l'estradiolo ha l'attività più alta, seguito da estrone ed estriolo (rapporto 1: 7: 100).
metabolismo degli estrogeni. Gli estrogeni circolano nel sangue in forma libera e legata alle proteine (biologicamente inattiva). Dal sangue, gli estrogeni entrano nel fegato, dove vengono inattivati dalla formazione di composti accoppiati con acido solforico e glucuronico, che entrano nei reni e vengono escreti nelle urine.
L'effetto degli estrogeni sul corpo si realizza come segue:
Effetto vegetativo (rigorosamente specifico) - gli estrogeni hanno un effetto specifico sugli organi genitali femminili: stimolano lo sviluppo dei caratteri sessuali secondari, causano iperplasia e ipertrofia dell'endometrio e del miometrio, migliorano l'afflusso di sangue all'utero, promuovono lo sviluppo del sistema escretore delle ghiandole mammarie;
- effetto generativo (meno specifico) - gli estrogeni stimolano i processi trofici durante la maturazione del follicolo, favoriscono la formazione e la crescita della granulosa, la formazione di un uovo e lo sviluppo del corpo luteo - preparano l'ovaio agli effetti degli ormoni gonadotropici;
- impatto complessivo(non specifico) - gli estrogeni in quantità fisiologica stimolano il sistema reticoloendoteliale (migliorano la produzione di anticorpi e l'attività dei fagociti, aumentando la resistenza dell'organismo alle infezioni), trattengono azoto, sodio, liquidi nei tessuti molli, calcio, fosforo nelle ossa . Provocano un aumento della concentrazione di glicogeno, glucosio, fosforo, creatinina, ferro e rame nel sangue e nei muscoli; ridurre il contenuto di colesterolo, fosfolipidi e grassi totali nel fegato e nel sangue, accelerare la sintesi degli acidi grassi superiori.
I gestageni sono secreti dalle cellule luteiniche del corpo luteo, dalle cellule luteinizzanti della granulosa e delle membrane follicolari, nonché dalla corteccia surrenale e dalla placenta. Il principale progestinico delle ovaie è il progesterone. Oltre al progesterone, le ovaie sintetizzano 17a-idrossiprogesterone, D4-pregnenol-20a-OH-3, D4-pregnenol-20b-OH-3.
Effetti dei gestageni:
Effetto vegetativo - i gestageni colpiscono i genitali dopo la stimolazione estrogenica preliminare: sopprimono la proliferazione dell'endometrio causata dagli estrogeni, svolgono trasformazioni secretorie nell'endometrio; durante la fecondazione dell'uovo, i gestageni sopprimono l'ovulazione, prevengono la contrazione uterina ("protettore" della gravidanza), promuovono lo sviluppo degli alveoli nelle ghiandole mammarie;
- effetto generativo - i gestageni a piccole dosi stimolano la secrezione di FSH, a grandi dosi bloccano sia l'FSH che l'LH; causare l'eccitazione del centro termoregolatore localizzato nell'ipotalamo, che si manifesta con un aumento della temperatura basale;
- effetto generale - gestagene in condizioni fisiologiche ridurre il contenuto di azoto amminico nel plasma sanguigno, aumentare l'escrezione di aminoacidi, migliorare la separazione succo gastrico inibire la secrezione della bile.
Gli androgeni sono secreti dalle cellule del rivestimento interno del follicolo, dalle cellule interstiziali (in piccola quantità) e dalle cellule della zona reticolare della corteccia surrenale (la fonte principale). I principali androgeni ovarici sono androstenedione e deidroepiandrosterone; testosterone ed epitestosterone sono sintetizzati a piccole dosi.
L'effetto specifico degli androgeni sul sistema riproduttivo dipende dal livello della loro secrezione (piccole dosi stimolano la funzione dell'ipofisi, grandi dosi la bloccano) e può manifestarsi come i seguenti effetti:
Effetto virile - grandi dosi gli androgeni causano l'ipertrofia del clitoride, la crescita dei capelli lungo tipo maschile, proliferazione della cartilagine cricoidea, comparsa di acne;
- effetto gonadotropico - piccole dosi di androgeni stimolano la secrezione di ormoni gonadotropici, favoriscono la crescita e la maturazione del follicolo, l'ovulazione, la luteinizzazione;
- effetto antigonadotropico - un alto livello di concentrazione di androgeni nel periodo preovulatorio sopprime l'ovulazione e successivamente provoca atresia follicolare;
- effetto estrogenico- a piccole dosi, gli androgeni provocano la proliferazione dell'endometrio e dell'epitelio vaginale;
- effetto antiestrogenico - grandi dosi di androgeni bloccano i processi di proliferazione nell'endometrio e portano alla scomparsa delle cellule acidofile nello striscio vaginale.
- effetto generale - gli androgeni hanno un'attività anabolica pronunciata, migliorano la sintesi proteica da parte dei tessuti; trattenere azoto, sodio e cloro nel corpo, ridurre l'escrezione di urea. Accelera la crescita ossea e l'ossificazione della cartilagine epifisaria, aumenta il numero dei globuli rossi e dell'emoglobina.
Altri ormoni ovarici: l'inibina, sintetizzata dalle cellule granulari, ha un effetto inibitorio sulla sintesi di FSH; ossitocina (trovata nel liquido follicolare, corpo luteo) - nelle ovaie ha un effetto luteolitico, favorisce la regressione del corpo luteo; la relaxina, formata nelle cellule della granulosa e del corpo luteo, favorisce l'ovulazione, rilassa il miometrio.
Utero
Sotto l'influenza degli ormoni ovarici, si osservano cambiamenti ciclici nel miometrio e nell'endometrio, corrispondenti alle fasi follicolina e luteale nelle ovaie. La fase follicolare è caratterizzata dall'ipertrofia delle cellule dello strato muscolare dell'utero, per la fase luteale - la loro iperplasia. I cambiamenti funzionali nell'endometrio si riflettono in un successivo cambiamento nelle fasi di rigenerazione, proliferazione, secrezione, desquamazione (mestruazioni).
La fase di rigenerazione (3-4 giorni del ciclo mestruale) è breve, caratterizzata dalla rigenerazione dell'endometrio dalle cellule dello strato basale.
L'epitelizzazione della superficie della ferita avviene dalle sezioni marginali delle ghiandole dello strato basale, nonché dalle sezioni profonde dello strato funzionale che non si espandono.
La fase di proliferazione (corrispondente alla fase follicolare) è caratterizzata da trasformazioni che avvengono sotto l'influenza degli estrogeni.
Fase iniziale della proliferazione (fino a 7-8 giorni del ciclo mestruale): la superficie della membrana mucosa è rivestita da un epitelio cilindrico appiattito, le ghiandole sembrano tubi corti dritti o leggermente contorti con un lume stretto, l'epitelio del le ghiandole sono a fila singola, basse, cilindriche.
La fase intermedia della proliferazione (fino a 10-12 giorni del ciclo mestruale): la superficie della mucosa è rivestita di epitelio prismatico, le ghiandole si allungano, diventano più tortuose, lo stroma è edematoso, allentato.
Fase avanzata della proliferazione (prima dell'ovulazione): le ghiandole diventano fortemente contorte, a volte a forma di sperone, il loro lume si espande, l'epitelio che riveste le ghiandole è multifilare, lo stroma è succoso, le arterie spirali raggiungono la superficie dell'endometrio, moderatamente contorto.
La fase di secrezione (corrispondente alla fase luteale) riflette i cambiamenti causati dall'azione del progesterone.
La fase iniziale della secrezione (prima del 18° giorno del ciclo mestruale) è caratterizzata dall'ulteriore sviluppo delle ghiandole e dall'espansione del loro lume, la maggior parte caratteristica questa fase - la comparsa nell'epitelio di vacuoli subnucleari contenenti glicogeno.
La fase intermedia della secrezione (19-23 giorni del ciclo mestruale) - riflette le trasformazioni caratteristiche del periodo di massimo splendore del corpo luteo, ad es. periodo di massima saturazione gestagena. Lo strato funzionale diventa più alto, chiaramente diviso in strati profondi e superficiali: profondo - spugnoso, spugnoso; superficiale - compatto. Le glandule si espandono, le loro pareti si piegano; un segreto contenente glicogeno e mucopolisaccaridi acidi appare nel lume delle ghiandole. Le arterie a spirale sono bruscamente tortuose, formano "palle" (il segno più affidabile che determina l'effetto luteinizzante). La struttura e lo stato funzionale dell'endometrio nei giorni 20-22 del ciclo mestruale di 28 giorni rappresentano le condizioni ottimali per l'impianto di blastocisti.
Fase tardiva della secrezione (24-27 giorni del ciclo mestruale): ci sono processi associati alla regressione del corpo luteo e, di conseguenza, una diminuzione della concentrazione degli ormoni da esso prodotti - il trofismo dell'endometrio è disturbato, il suo si formano alterazioni degenerative.
Morfologicamente, l'endometrio regredisce, compaiono i segni della sua ischemia. Ciò riduce la succosità del tessuto, che porta all'increspatura dello stroma dello strato funzionale. La piegatura delle pareti delle ghiandole è intensificata. Il 26-27° giorno del ciclo mestruale si osservano dilatazioni lacunari dei capillari ed emorragie focali nello stroma nelle zone superficiali dello strato compatto; a causa dello scioglimento delle strutture fibrose, compaiono aree di separazione delle cellule dello stroma e dell'epitelio delle ghiandole. Questa condizione dell'endometrio viene definita "mestruazione anatomica" e precede immediatamente le mestruazioni cliniche.
Fase emorragica, desquamazione (28-29 giorni del ciclo mestruale). Nel meccanismo del sanguinamento mestruale, il ruolo di primo piano è dato ai disturbi circolatori causati da spasmi prolungati delle arterie (stasi, formazione di trombi, fragilità e permeabilità della parete vascolare, emorragie nello stroma, infiltrazione leucocitaria). Il risultato di queste trasformazioni sono la necrobiosi tissutale e il suo scioglimento. A causa dell'espansione dei vasi sanguigni che si verifica dopo un lungo spasmo, una grande quantità di sangue entra nel tessuto endometriale, che porta alla rottura dei vasi e al rigetto - desquamazione - delle sezioni necrotiche dello strato funzionale dell'endometrio, ad es. al sanguinamento mestruale.
I tessuti bersaglio sono i punti di applicazione dell'azione degli ormoni sessuali. Questi includono: tessuto cerebrale, organi riproduttivi, ghiandole mammarie, follicoli piliferi e pelle, ossa, tessuto adiposo. Le cellule di questi organi e tessuti contengono recettori per gli ormoni sessuali. Il mediatore di questo livello di regolazione del sistema riproduttivo è il cAMP, che regola il metabolismo nelle cellule dei tessuti bersaglio in base alle esigenze dell'organismo in risposta agli effetti degli ormoni. I regolatori intercellulari includono anche le prostaglandine, che sono formate da acidi grassi insaturi in tutti i tessuti del corpo. L'azione delle prostaglandine si realizza attraverso cAMP.
Il cervello è l'organo bersaglio degli ormoni sessuali. Gli ormoni sessuali attraverso i fattori di crescita possono influenzare sia i neuroni che le cellule gliali. Gli ormoni sessuali influenzano la formazione di segnali in quelle aree del SNC che sono coinvolte nella regolazione del comportamento riproduttivo (nuclei ventromediali, ipotalamici e dell'amigdala), così come nelle aree che regolano la sintesi e il rilascio di ormoni da parte della ghiandola pituitaria (in nucleo arcuato ipotalamico e nell'area preottica).
Nell'ipotalamo, l'obiettivo principale degli ormoni sessuali sono i neuroni che formano il nucleo arcuato, in cui il GnRH viene sintetizzato e rilasciato in modalità pulsata. Gli oppioidi possono avere un effetto eccitatorio e inibitorio sui neuroni che sintetizzano il GnRH nell'ipotalamo. Gli estrogeni stimolano la sintesi dei recettori per gli oppioidi endogeni. La β-endorfina (β-EF) è il peptide oppioide endogeno più attivo che influenza il comportamento, provoca analgesia, è coinvolto nella termoregolazione e ha proprietà neuroendocrine. Nella postmenopausa e dopo l'ovariectomia, il livello di r-EF diminuisce, il che contribuisce al verificarsi di vampate di calore e sudorazione eccessiva, nonché a cambiamenti dell'umore, del comportamento e dei disturbi monicettivi. Gli estrogeni eccitano il SNC aumentando la sensibilità dei recettori dei neurotrasmettitori nei neuroni sensibili agli estrogeni, portando a un aumento dell'umore, una maggiore attività e effetti antidepressivi. Bassi livelli di estrogeni in menopausa causano lo sviluppo della depressione.
Gli androgeni svolgono anche un ruolo nel comportamento sessuale, nelle risposte emotive e nella funzione cognitiva di una donna. La carenza di androgeni in menopausa porta ad una diminuzione dei peli pubici, della forza muscolare e di una diminuzione della libido.
Le tube di Falloppio
Lo stato funzionale delle tube di Falloppio varia a seconda della fase del ciclo mestruale. Quindi, nella fase luteale del ciclo, viene attivato l'apparato ciliato dell'epitelio ciliato, l'altezza delle sue cellule aumenta, sulla parte apicale di cui si accumula un segreto. Anche il tono dello strato muscolare dei tubi cambia: al momento dell'ovulazione si registra una diminuzione e un'intensificazione delle loro contrazioni, che hanno sia un carattere pendolare che rotazionale-traslazionale. L'attività muscolare è diseguale in diverse parti dell'organo: le onde peristaltiche sono più caratteristiche delle parti distali. Attivazione dell'apparato ciliato dell'epitelio ciliato, labilità tono muscolare le tube di Falloppio nella fase luteale, l'asincronismo e l'eterogeneità dell'attività contrattile in diverse parti dell'organo sono determinati collettivamente per garantire condizioni ottimali per il trasporto dei gameti.
Inoltre, in diverse fasi del ciclo mestruale, cambia la natura della microcircolazione nei vasi delle tube di Falloppio. Nel periodo dell'ovulazione, le vene che circondano l'imbuto e penetrano in profondità nelle sue frange traboccano di sangue, per cui il tono della fimbria aumenta e l'imbuto, avvicinandosi all'ovaio, lo copre, che, parallelamente ad altri meccanismi, assicura che l'uovo ovulato entri nel tubo. Quando cessa il ristagno di sangue nelle vene anulari dell'imbuto, quest'ultimo si allontana dalla superficie dell'ovaio.
Vagina
Durante il ciclo mestruale, la struttura dell'epitelio vaginale subisce fasi proliferative e regressive. La fase proliferativa corrisponde allo stadio follicolinico delle ovaie ed è caratterizzata da crescita, ingrossamento e differenziazione delle cellule epiteliali. Durante il periodo corrispondente alla fase iniziale della follicolina, la crescita dell'epitelio avviene principalmente a causa delle cellule dello strato basale; a metà della fase, il contenuto di cellule intermedie aumenta. Nel periodo preovulatorio, quando l'epitelio vaginale raggiunge il suo massimo spessore - 150-300 micron - si ha un'attivazione della maturazione delle cellule dello strato superficiale.
La fase regressiva corrisponde allo stadio luteale. In questa fase, la crescita dell'epitelio si interrompe, il suo spessore diminuisce, alcune cellule subiscono uno sviluppo inverso. La fase termina con la desquamazione delle cellule in gruppi grandi e compatti.
Le ghiandole mammarie aumentano durante il ciclo mestruale, a partire dal momento dell'ovulazione e raggiungendo il massimo entro il primo giorno delle mestruazioni. Prima delle mestruazioni, c'è un aumento del flusso sanguigno, un aumento del contenuto di liquidi nel tessuto connettivo, lo sviluppo di edema interlobulare e l'espansione dei dotti interlobulari, che porta ad un aumento della ghiandola mammaria.
Regolazione neuroumorale del ciclo mestruale
La regolazione del normale ciclo mestruale viene effettuata a livello di neuroni cerebrali specializzati che ricevono informazioni sullo stato dell'ambiente interno ed esterno e le convertono in segnali neuroormonali. Questi ultimi attraverso il sistema dei neurotrasmettitori entrano nelle cellule neurosecretorie dell'ipotalamo e stimolano la secrezione di GnRH. Il GnRH attraverso la rete circolatoria locale del sistema portale ipotalamo-ipofisario penetra direttamente nell'adenoipofisi, dove provvede alla secrezione circolare e al rilascio di gonadotropine glicoproteiche: FSH e LH. Entrano nelle ovaie sistema circolatorio: FSH stimola la crescita e la maturazione del follicolo, LH - steroidogenesi. Sotto l'influenza di FSH e LH, le ovaie producono estrogeni e progesterone con la partecipazione di PRL, che, a loro volta, causano trasformazioni cicliche negli organi bersaglio: l'utero, le tube di Falloppio, la vagina, nonché nella pelle, follicoli piliferi, ossa, tessuto adiposo, cervello.
Lo stato funzionale del sistema riproduttivo è regolato da alcuni collegamenti tra i suoi sottosistemi costitutivi:
a) un lungo anello tra le ovaie ei nuclei dell'ipotalamo;
b) un lungo anello tra gli ormoni delle ovaie e la ghiandola pituitaria;
c) un circuito ultracorto tra l'ormone di rilascio delle gonadotropine ei neurociti ipotalamici.
La relazione tra questi sottosistemi si basa sul principio del feedback, che ha sia un carattere negativo (interazione più-meno) che positivo (interazione più-più). L'armonia dei processi che si verificano nel sistema riproduttivo è determinata da: l'utilità della stimolazione gonadotropica; il normale funzionamento delle ovaie, in particolare il corretto flusso dei processi nella vescicola di Graaf e nel corpo luteo che si forma poi al suo posto; la corretta interazione dei collegamenti periferici e centrali - afferentazione inversa.
Il ruolo delle prostaglandine nella regolazione del sistema riproduttivo femminile
Le prostaglandine rappresentano una classe speciale di sostanze biologicamente attive (acidi grassi idrossilati insaturi) che si trovano in quasi tutti i tessuti del corpo. Le prostaglandine sono sintetizzate all'interno della cellula e rilasciate nelle stesse cellule su cui agiscono. Pertanto, le prostaglandine sono chiamate ormoni cellulari. Non vi è alcuna scorta di prostaglandine nel corpo umano, poiché vengono inattivate in un breve periodo quando entrano nel flusso sanguigno. Gli estrogeni e l'ossitocina migliorano la sintesi delle prostaglandine, il progesterone e la prolattina hanno un effetto inibitorio. I farmaci antinfiammatori non steroidei hanno un potente effetto antiprostaglandine.
Il ruolo delle prostaglandine nella regolazione del sistema riproduttivo femminile:
1. Partecipazione al processo di ovulazione. Sotto l'influenza degli estrogeni, il contenuto di prostaglandine nelle cellule della granulosa raggiunge il massimo al momento dell'ovulazione e fornisce una rottura della parete follicolare matura (le prostaglandine aumentano l'attività contrattile degli elementi muscolari lisci del guscio follicolare e riducono la formazione di collagene). Le prostaglandine sono anche attribuite alla capacità di luteolisi - regressione del corpo luteo.
2. Trasporto dell'uovo. Le prostaglandine influenzano l'attività contrattile delle tube di Falloppio: nella fase follicolare provocano la contrazione della sezione istmica delle tube, nella fase luteale - il suo rilassamento, aumento della peristalsi dell'ampolla, che contribuisce alla penetrazione dell'uovo nell'utero cavità. Inoltre, le prostaglandine agiscono sul miometrio: dagli angoli delle tube verso il fondo uterino, l'effetto stimolante delle prostaglandine viene sostituito da uno inibitorio e, quindi, favorisce la nidazione della blastocisti.
3. Regolazione del sanguinamento mestruale. L'intensità delle mestruazioni è determinata non solo dalla struttura dell'endometrio al momento del suo rigetto, ma anche dall'attività contrattile del miometrio, dalle arteriole e dall'aggregazione piastrinica.
Questi processi sono strettamente correlati al grado di sintesi e degradazione delle prostaglandine.
Ciclo mestruale- cambiamenti ciclici nel corpo di una donna, specialmente nel sistema riproduttivo, la cui manifestazione esterna è sanguinamento dal tratto genitale - mestruazioni. Il ciclo mestruale si instaura dopo il menarca (prima mestruazione) e persiste per tutto il periodo riproduttivo, o fertile, della vita di una donna con la capacità di riprodurre la prole. I cambiamenti ciclici nel corpo di una donna sono bifasici. La prima fase (follicolina) del ciclo è determinata dalla maturazione del follicolo e dell'uovo nell'ovaio, dopo di che si rompe e l'uovo lo lascia - l'ovulazione. La seconda fase (luteale) è associata alla formazione del corpo luteo.
Allo stesso tempo, in modo ciclico, la rigenerazione e la proliferazione dello strato funzionale avvengono in sequenza nell'endometrio, che è sostituito dall'attività secretoria delle sue ghiandole. I cambiamenti nell'endometrio terminano con la desquamazione dello strato funzionale (mestruazioni). Il significato biologico dei cambiamenti che si verificano durante il ciclo mestruale nelle ovaie e nell'endometrio è quello di garantire la funzione riproduttiva nelle fasi di maturazione dell'uovo, la sua fecondazione e l'impianto dell'embrione nell'utero. Se non si verifica la fecondazione dell'uovo, lo strato funzionale dell'endometrio viene respinto, compare una secrezione sanguinolenta dal tratto genitale e si verificano nuovamente e nella stessa sequenza nel sistema riproduttivo processi volti a garantire la maturazione dell'uovo.
Mestruazioni- questa è una secrezione sanguinolenta dal tratto genitale che si ripete a determinati intervalli durante l'intero periodo riproduttivo della vita di una donna al di fuori della gravidanza e dell'allattamento. Le mestruazioni sono il culmine del ciclo mestruale e si verificano alla fine della sua fase luteale a causa del rigetto dello strato funzionale dell'endometrio. La prima mestruazione (menarhe) si verifica all'età di 10-12 anni. Nei successivi 1 - 1,5 anni, le mestruazioni possono essere irregolari e solo allora viene stabilito un ciclo mestruale regolare. Il primo giorno delle mestruazioni viene considerato condizionatamente come primo giorno del ciclo e la durata del ciclo viene calcolata come l'intervallo tra i primi giorni di due mestruazioni successive.
ginecologia dell'ovulazione riproduttiva mestruale
Riso. uno. Regolazione ormonale del ciclo mestruale (schema): a - il cervello; b - cambiamenti nell'ovaio; c - cambiamento nel livello degli ormoni; d - cambiamenti nell'endometrio
Parametri esterni del normale ciclo mestruale: durata da 21 a 35 giorni (per il 60% delle donne la durata media del ciclo è di 28 giorni); durata del flusso mestruale da 2 a 7 giorni; la quantità di perdita di sangue nei giorni mestruali 40-60 ml (in media 50 ml).
I processi che assicurano il normale svolgimento del ciclo mestruale sono regolati da un unico sistema neuroendocrino funzionale, che comprende reparti centrali (integrative) e strutture periferiche (effettore) con un certo numero di collegamenti intermedi. In accordo con la loro gerarchia (dalle più alte strutture regolamentari al diretto organi esecutivi) nella regolazione neuroendocrina si possono distinguere 5 livelli, che interagiscono secondo il principio delle relazioni positive e negative dirette e inverse (Fig.)
Il primo (più alto) livello di regolamentazione il funzionamento del sistema riproduttivo sono le strutture che costituiscono l'accettore di tutte le influenze esterne e interne (dai reparti subordinati): la corteccia cerebrale del sistema nervoso centrale e le strutture cerebrali extraipotalamiche (sistema limbico, ippocampo, amigdala). Adeguatezza della percezione del SNC influenze esterne e, di conseguenza, la sua influenza sui dipartimenti subordinati che regolano i processi nel sistema riproduttivo dipende dalla natura degli stimoli esterni (forza, frequenza e durata della loro azione), nonché dallo stato iniziale del SNC, che colpisce la sua resistenza ai carichi di sollecitazione.
È ben noto la possibilità di interrompere le mestruazioni in condizioni di forte stress (perdita di persone care, condizioni di guerra, ecc.), nonché senza evidenti influenze esterne con squilibrio mentale generale ("falsa gravidanza" - ritardo delle mestruazioni con un forte desiderio o con una forte paura rimanere incinta). I reparti regolatori superiori dell'apparato riproduttivo percepiscono le influenze interne attraverso specifici recettori per i principali ormoni sessuali: estrogeni, progesterone e androgeni. In risposta a stimoli esterni ed interni nella corteccia cerebrale e nelle strutture extraipotalamiche, si verificano la sintesi, il rilascio e il metabolismo di neuropeptidi, neurotrasmettitori, nonché la formazione di recettori specifici, che a loro volta influenzano selettivamente la sintesi e il rilascio dell'ormone di rilascio di l'ipotalamo. Ai neurotrasmettitori più importanti, es. I trasmettitori includono noradrenalina, dopamina, acido gamma-aminobutirrico (GABA), acetilcolina, serotonina e melatonina. I neurotrasmettitori cerebrali regolano la produzione dell'ormone di rilascio delle gonadotropine (GnRH): noradrenalina, acetilcolina e GABA ne stimolano il rilascio, mentre la dopamina e la serotonina hanno l'effetto opposto.
Neuropeptidi(peptidi oppioidi endogeni - EOP, fattore di rilascio della corticotropina e galanina) influenzano anche la funzione dell'ipotalamo e l'equilibrio del funzionamento di tutte le parti del sistema riproduttivo. Attualmente, ci sono 3 gruppi di EOP: encefaline, endorfine e dinorfine. Queste sostanze si trovano non solo in varie strutture del cervello e del sistema nervoso autonomo, ma anche nel fegato, nei polmoni, nel pancreas e in altri organi, nonché in alcuni fluidi biologici (plasma sanguigno, contenuto del follicolo). Secondo i concetti moderni, le EOP sono coinvolte nella regolazione della formazione del GnRH. Un aumento del livello di EOP sopprime la secrezione di GnRH e, di conseguenza, il rilascio di LH e FSH, che possono essere la causa dell'anovulazione, e altro casi gravi- amenorrea. È con un aumento dell'EOP che l'insorgenza di varie forme di amenorrea della genesi centrale è associata allo stress, nonché all'eccessivo sforzo fisico, ad esempio negli atleti. La nomina di inibitori del recettore degli oppioidi (farmaci come il naloxone) normalizza la formazione di GnRH, che contribuisce alla normalizzazione della funzione ovulatoria e di altri processi nel sistema riproduttivo nei pazienti con amenorrea centrale. Con una diminuzione del livello di steroidi sessuali (con arresto della funzione ovarica correlato all'età o chirurgico), l'EOP non ha un effetto inibitorio sul rilascio di GnRH, che probabilmente causa un aumento della produzione di gonadotropine nelle donne in postmenopausa. Pertanto, l'equilibrio della sintesi e le successive trasformazioni metaboliche di neurotrasmettitori, neuropeptidi e neuromodulatori nei neuroni cerebrali e nelle strutture sovraipotalamiche assicura il normale corso dei processi associati alla funzione ovulatoria e mestruale.
Il secondo livello di regolamentazione la funzione riproduttiva è l'ipotalamo, in particolare la sua zona pituitaria, costituita dai neuroni dei nuclei arcuati ventro e dorsomediale, che hanno attività neurosecretoria. Queste cellule hanno le proprietà sia dei neuroni (riproduzione degli impulsi elettrici regolatori) che delle cellule endocrine, che hanno un effetto stimolante (liberina) o bloccante (statina). L'attività di neurosecrezione nell'ipotalamo è regolata sia dagli ormoni sessuali che provengono dal flusso sanguigno sia dai neurotrasmettitori e neuropeptidi formati nella corteccia cerebrale e nelle strutture sovraipotalamiche. L'ipotalamo secerne il GnRH, che contiene gli ormoni follicolo-stimolanti (RGFSH - folliberina) e luteinizzante (RGLG - luliberina), che agiscono sulla ghiandola pituitaria. L'ormone di rilascio LH (RGLG - luliberin) è stato isolato, sintetizzato e descritto in dettaglio. Ad oggi, non è stato possibile isolare e sintetizzare l'ormone follicolo-stimolante di rilascio. Tuttavia, è stato stabilito che il decapeptide RGLG e i suoi analoghi sintetici stimolano il rilascio di gonadi da parte degli otrofi non solo di LH, ma anche di FSH. A questo proposito, è stato adottato un termine per i liberins gonadotropici - l'ormone di rilascio delle gonadotropine (GnRH), che è essenzialmente un sinonimo di RHLH. Anche la liberina ipotalamica, che stimola la formazione di prolattina, non è stata identificata, sebbene sia stato stabilito che la sua sintesi è attivata dall'ormone di rilascio del TSH (tiroliberina). La formazione della prolattina è attivata anche dalla serotonina e dai peptidi oppioidi endogeni che stimolano i sistemi serotoninergici. La dopamina, al contrario, inibisce il rilascio di prolattina dai lattotrofi dell'adenoipofisi. L'uso di farmaci dopaminergici come il parlodel (bromkriptin) può trattare con successo l'iperprolattinemia funzionale e organica, che è una causa molto comune di disturbi mestruali e ovulatori. La secrezione di GnRH è programmata geneticamente e ha un carattere pulsatile (circolare): picchi di aumento della secrezione ormonale della durata di diversi minuti sono sostituiti da intervalli di 1-3 ore di attività secretoria relativamente bassa. La frequenza e l'ampiezza della secrezione di GnRH regolano il livello di estradiolo: le emissioni di GnRH nel periodo preovulatorio sullo sfondo del rilascio massimo di estradiolo sono significativamente maggiori rispetto alle prime fasi follicolare e luteale.
Il terzo livello di regolamentazione la funzione riproduttiva è la ghiandola pituitaria anteriore, in cui vengono secreti gli ormoni gonadotropi - follicolo-stimolanti o follitropina (FSH) e luteinizzante o lutropina (LH), prolattina, ormone adrenocorticotropo (ACTH), ormone somatotropo (STH) e tiroide- ormone stimolante (TSH). Il normale funzionamento del sistema riproduttivo è possibile solo con una selezione equilibrata di ciascuno di essi. L'FSH stimola la crescita e la maturazione dei follicoli nell'ovaio, la proliferazione delle cellule della granulosa; la formazione dei recettori FSH e LH sulle cellule della granulosa; attività dell'aromatasi nel follicolo in maturazione (questo migliora la conversione degli androgeni in estrogeni); produzione di inibina, attivina e fattori di crescita simili all'insulina. LH promuove la formazione di androgeni nelle cellule della teca; ovulazione (insieme a FSH); rimodellamento delle cellule della granulosa durante la luteinizzazione; sintesi del progesterone nel corpo luteo. La prolattina ha una varietà di effetti sul corpo di una donna. Il suo principale ruolo biologico- stimolazione della crescita delle ghiandole mammarie, regolazione dell'allattamento e controllo della secrezione di progesterone da parte del corpo luteo attivando la formazione di recettori per LH in esso Durante la gravidanza e l'allattamento, l'inibizione della sintesi della prolattina si interrompe e, di conseguenza, un aumento del suo livello nel sangue.
Al quarto livello la regolazione della funzione riproduttiva include la periferica organi endocrini(ovaie, ghiandole surrenali, tiroide). Il ruolo principale appartiene alle ovaie e altre ghiandole svolgono le proprie funzioni specifiche, pur mantenendo il normale funzionamento del sistema riproduttivo. Nelle ovaie si verificano la crescita e la maturazione dei follicoli, l'ovulazione, la formazione del corpo luteo e la sintesi degli steroidi sessuali. Alla nascita, le ovaie di una ragazza contengono circa 2 milioni di follicoli primordiali. La maggior parte di loro subisce cambiamenti atretici per tutta la vita e solo una piccolissima parte attraversa un ciclo di sviluppo completo dal primordiale alla maturazione con la successiva formazione del corpo luteo. Al momento del menarca, le ovaie contengono 200-400 mila follicoli primordiali. Durante un ciclo mestruale, di regola, si sviluppa solo un follicolo con un uovo all'interno. In caso di maturazione di un numero maggiore, è possibile la gravidanza multipla.
Follicogenesi inizia sotto l'influenza dell'FSH nella parte avanzata della fase luteale del ciclo e termina all'inizio del picco di rilascio delle gonadotropine. Circa 1 giorno prima dell'inizio delle mestruazioni, il livello di FSH aumenta nuovamente, il che garantisce l'ingresso in crescita, o reclutamento, dei follicoli (1-4° giorno del ciclo), selezione del follicolo da una coorte omogenea - quasi -sincronizzato (5--7° giorno), maturazione del follicolo dominante (8-12° giorno) e ovulazione (13-15° giorno). Questo processo, che costituisce la fase follicolare, dura circa 14 giorni. Di conseguenza, si forma un follicolo preovulatorio e il resto della coorte di follicoli che sono entrati in crescita subisce atresia. La selezione di un singolo follicolo destinato all'ovulazione è inseparabile dalla sintesi degli estrogeni in esso contenuti. La stabilità della produzione di estrogeni dipende dall'interazione tra teca e cellule della granulosa, la cui attività, a sua volta, è modulata da numerosi meccanismi endocrini, paracrini e autocrini che regolano la crescita e la maturazione dei follicoli. A seconda dello stadio di sviluppo e delle caratteristiche morfologiche, si distinguono i follicoli primordiali, preantrali, antrali e preovulatori o dominanti. Il follicolo primordiale è costituito da un ovulo immaturo, che si trova nell'epitelio follicolare e granulare (granulare). All'esterno, il follicolo è circondato da una membrana di tessuto connettivo (cellule della teca). Durante ogni ciclo mestruale, iniziano a crescere da 3 a 30 follicoli primordiali, trasformandosi in follicoli preantrali (primari). follicolo preantrale. Nel follicolo preantrale, l'ovocita aumenta di dimensioni ed è circondato da una membrana chiamata zona pellucida. Le cellule epiteliali della granulosa proliferano e si arrotondano, formando uno strato granulare del follicolo (stratum granulosum) e lo strato di teca è formato dallo stroma circostante. Questa fase è caratterizzata dall'attivazione della produzione di estrogeni formati nello strato di granulosa.
Follicolo preovulatoria (dominante).(Fig. 2.2) spicca tra i follicoli in crescita per le dimensioni maggiori (il diametro al momento dell'ovulazione raggiunge i 20 mm). Il follicolo dominante ha uno strato riccamente vascolarizzato di cellule della teca e cellule della granulosa con un gran numero di recettori per FSH e LH. Insieme alla crescita e allo sviluppo del follicolo preovulatorio dominante nelle ovaie, l'atresia dei follicoli rimanenti (reclutati) che inizialmente sono entrati nella crescita si verifica in parallelo e continua anche l'atresia dei follicoli primordiali. Durante la maturazione, nel follicolo preovulatore si verifica un aumento di 100 volte del volume del liquido follicolare. Nel processo di maturazione dei follicoli antrali, la composizione del liquido follicolare cambia.
Follicolo antrale (secondario). subisce un aumento della cavità formata dall'accumulo di liquido follicolare prodotto dalle cellule dello strato granuloso. Aumenta anche l'attività di formazione di steroidi sessuali. Le cellule della teca sintetizzano gli androgeni (androstenedione e testosterone). Una volta nelle cellule della granulosa, gli androgeni subiscono attivamente l'aromatizzazione, che determina la loro conversione in estrogeni. In tutte le fasi dello sviluppo del follicolo, ad eccezione del preovulatore, il contenuto di progesterone è a un livello costante e relativamente basso. Le gonadotropine e la prolattina nel liquido follicolare sono sempre inferiori a quelle nel plasma sanguigno e il livello di prolattina tende a diminuire con la maturazione del follicolo. L'FSH è determinato dall'inizio della formazione della cavità e l'LH può essere rilevato solo in un follicolo preovulatorio maturo insieme al progesterone. Il liquido follicolare contiene anche ossitocina e vasopressina e in concentrazioni 30 volte superiori a quelle nel sangue, che possono indicare la formazione locale di questi neuropeptidi. Le prostaglandine delle classi E ed F vengono rilevate solo nel follicolo preovulatore e solo dopo l'inizio dell'aumento del livello di LH, che indica il loro coinvolgimento diretto nel processo di ovulazione.
Ovulazione- rottura del follicolo preovulatorio (dominante) e rilascio dell'uovo da esso. L'ovulazione è accompagnata da sanguinamento dai capillari distrutti che circondano le cellule della teca (Fig. 2.3). Si ritiene che l'ovulazione avvenga 24-36 ore dopo il picco preovulatoria dell'estradiolo, che provoca un forte aumento della secrezione di LH. In questo contesto, vengono attivati gli enzimi proteolitici - collagenasi e plasmina, che distruggono il collagene della parete follicolare e quindi ne riducono la forza. Allo stesso tempo, l'aumento osservato della concentrazione di prostaglandina F2a, così come l'ossitocina, induce la rottura del follicolo a causa della loro stimolazione della contrazione della muscolatura liscia e dell'espulsione dell'ovocita con il tumulo oviposito dalla cavità del follicolo . La rottura del follicolo è facilitata anche da un aumento della concentrazione di prostaglandina E2 e relaxina in essa contenuta, che riducono la rigidità delle sue pareti. Dopo il rilascio dell'uovo, i capillari risultanti crescono rapidamente nella cavità del follicolo ovulato. Le cellule della granulosa subiscono luteinizzazione, manifestata morfologicamente in un aumento del loro volume e nella formazione di inclusioni lipidiche. Questo processo, che porta alla formazione del corpo luteo, è stimolato dall'LH, che interagisce attivamente con specifici recettori cellulari della granulosa.
corpo luteo- una formazione ormonale transitoria attiva, funzionante per 14 giorni, indipendentemente dalla durata totale del ciclo mestruale. Se la gravidanza non si verifica, il corpo luteo regredisce. Un corpo luteo completo si sviluppa solo nella fase in cui nel follicolo preovulatore si forma una quantità adeguata di cellule della granulosa ad alto contenuto di recettori LH.Nel periodo riproduttivo, le ovaie sono la principale fonte di estrogeni (estradiolo, estriolo ed estrone). , di cui l'estradiolo è il più attivo. Oltre agli estrogeni, nelle ovaie vengono prodotti il progesterone e una certa quantità di androgeni. Oltre agli ormoni steroidei e alle inibine che entrano nel flusso sanguigno e colpiscono gli organi bersaglio, nelle ovaie vengono sintetizzati anche composti biologicamente attivi con un effetto simil-ormonale prevalentemente locale. Pertanto, le prostaglandine formate, l'ossitocina e la vasopressina svolgono un ruolo importante come fattori scatenanti dell'ovulazione. L'ossitocina ha anche un effetto luteolitico, fornendo regressione del corpo luteo. La relaxina favorisce l'ovulazione e ha un effetto tocolitico sul miometrio. Fattori di crescita-- il fattore di crescita epidermico (EGF) ei fattori di crescita insulino-simili 1 e 2 (IPFR-1 e IPFR-2) attivano la proliferazione delle cellule della granulosa e la maturazione dei follicoli. Gli stessi fattori sono coinvolti insieme alle gonadotropine nella regolazione fine dei processi di selezione del follicolo dominante, nell'atresia dei follicoli degeneranti di tutti gli stadi, nonché nella cessazione del funzionamento del corpo luteo. La formazione di androgeni nelle ovaie rimane stabile durante tutto il ciclo. Lo scopo biologico principale della secrezione ciclica di steroidi sessuali nell'ovaio è la regolazione dei cambiamenti ciclici fisiologici nell'endometrio. Gli ormoni ovarici non determinano solo i cambiamenti funzionali nel sistema riproduttivo stesso. Inoltre influenzano attivamente i processi metabolici in altri organi e tessuti che hanno recettori per gli steroidi sessuali. Questi recettori possono essere citoplasmatici (recettori del citosol) o nucleari.
I recettori citoplasmatici sono strettamente specifici per estrogeni, progesterone e testosterone, mentre i recettori nucleari possono accettare non solo gli ormoni steroidei, ma anche aminopeptidi, insulina e glucagone. Per il legame con il recettore del progesterone, i glucocorticoidi sono considerati antagonisti. Nella pelle, sotto l'influenza dell'estradiolo e del testosterone, viene attivata la sintesi del collagene, che aiuta a mantenerne l'elasticità. Aumento del sebo, acne, follicolite, porosità e crescita eccessiva dei capelli associata ad una maggiore esposizione agli androgeni. Nelle ossa, estrogeni, progesterone e androgeni supportano il normale rimodellamento prevenendo il riassorbimento osseo. Nel tessuto adiposo, l'equilibrio di estrogeni e androgeni predetermina sia l'attività del suo metabolismo che la distribuzione nel corpo. Gli steroidi sessuali (progesterone) modulano significativamente il lavoro del centro di termoregolazione ipotalamico. Con i recettori per gli steroidi sessuali nel sistema nervoso centrale, nelle strutture dell'ippocampo che regolano la sfera emotiva, nonché nei centri che controllano le funzioni autonome, si associa il fenomeno dell'"onda mestruale" nei giorni precedenti le mestruazioni. Questo fenomeno si manifesta con uno squilibrio nei processi di attivazione e inibizione nella corteccia, fluttuazioni del tono dei sistemi simpatico e parasimpatico (influendo in modo particolare sul funzionamento del sistema cardiovascolare), nonché cambiamenti di umore e una certa irritabilità. Nelle donne sane, questi cambiamenti, tuttavia, non vanno oltre i confini fisiologici.
Quinto livello la regolazione della funzione riproduttiva è sensibile alle fluttuazioni dei livelli di steroidi sessuali nelle parti interne ed esterne del sistema riproduttivo (utero, tube di Falloppio, mucosa vaginale), così come le ghiandole mammarie. I cambiamenti ciclici più pronunciati si verificano nell'endometrio.
Cambiamenti ciclici nell'endometrio toccare il suo strato superficiale, costituito da cellule epiteliali compatte, e l'intermedio, che vengono respinte durante le mestruazioni. Lo strato basale, che non viene respinto durante le mestruazioni, assicura il ripristino degli strati desquamati. In base alle variazioni dell'endometrio durante il ciclo, si distinguono la fase di proliferazione, la fase di secrezione e la fase di sanguinamento (mestruazioni).
Fase di proliferazione("follicolare") dura in media 12-14 giorni, a partire dal 5° giorno del ciclo.Durante questo periodo, un nuovo strato superficiale con ghiandole tubulari allungate rivestite da epitelio colonnare con aumentata attività mitotica. Lo spessore dello strato funzionale dell'endometrio è di 8 mm.
Fase di secrezione (luteale) associato all'attività del corpo luteo, dura 14 giorni (+ 1 giorno). Durante questo periodo, l'epitelio delle ghiandole endometriali inizia a produrre un segreto contenente glicosaminoglicani acidi, glicoproteine e glicogeno. L'attività di secrezione diventa la più alta nel giorno 20-21. A questo punto, la quantità massima di enzimi proteolitici si trova nell'endometrio e nello stroma si verificano trasformazioni deciduali (le cellule dello strato compatto diventano più grandi, acquisendo una forma arrotondata o poligonale, il glicogeno si accumula nel loro citoplasma). C'è una forte vascolarizzazione dello stroma: le arterie a spirale sono fortemente tortuose, formano "grovigli" che si trovano nell'intero strato funzionale. Le vene sono dilatate. Tali cambiamenti nell'endometrio, osservati il 20-22° giorno (6-8° giorno dopo l'ovulazione) del ciclo mestruale di 28 giorni, forniscono le migliori condizioni per l'impianto di un ovulo fecondato. Entro il 24°-27° giorno, a causa dell'inizio della regressione del corpo luteo e della diminuzione della concentrazione di ormoni da esso prodotti, il trofismo dell'endometrio viene disturbato con un graduale aumento dei cambiamenti degenerativi in esso. Dalle cellule granulari dello stroma endometriale vengono rilasciati granuli contenenti relaxina, che prepara il rigetto mestruale della mucosa. Nelle aree superficiali dello strato compatto si notano dilatazioni lacunari dei capillari ed emorragie nello stroma, rilevabili in 1 giorno. prima dell'inizio delle mestruazioni.
Mestruazioni comprende la desquamazione e la rigenerazione dello strato funzionale dell'endometrio. A causa della regressione del corpo luteo e di una forte diminuzione del contenuto di steroidi sessuali nell'endometrio, l'ipossia aumenta. L'inizio delle mestruazioni è facilitato da uno spasmo prolungato delle arterie, che porta alla stasi del sangue e alla formazione di coaguli di sangue. L'ipossia tissutale (acidosi tissutale) è esacerbata da una maggiore permeabilità dell'endotelio, fragilità delle pareti dei vasi, numerose piccole emorragie e massiccia infiltrazione di leucociti. Gli enzimi proteolitici lisosomiali rilasciati dai leucociti favoriscono la fusione degli elementi tissutali. A seguito di uno spasmo prolungato dei vasi, la loro espansione paretica si verifica con un aumento del flusso sanguigno. Allo stesso tempo, si nota un aumento della pressione idrostatica nel microcircolo e una rottura delle pareti dei vasi, che a questo punto hanno in gran parte perso la loro resistenza meccanica. In questo contesto, si verifica la desquamazione attiva delle aree necrotiche dello strato funzionale. Entro la fine del 1° giorno delle mestruazioni, 2/3 dello strato funzionale viene respinto e la sua completa desquamazione di solito termina il 3° giorno. La rigenerazione dell'endometrio inizia immediatamente dopo il rigetto dello strato funzionale necrotico.
La base per la rigenerazione sono le cellule epiteliali dello stroma dello strato basale. In condizioni fisiologiche, già al 4° giorno del ciclo, l'intera superficie della ferita della mucosa viene riepitelizzata. Questo è seguito di nuovo da cambiamenti ciclici nell'endometrio: le fasi di proliferazione e secrezione. Cambiamenti successivi durante il ciclo nell'endometrio: la proliferazione, la secrezione e le mestruazioni dipendono non solo dalle fluttuazioni cicliche dei livelli di steroidi sessuali nel sangue, ma anche dallo stato dei recettori tissutali di questi ormoni. La concentrazione dei recettori nucleari dell'estradiolo aumenta fino alla metà del ciclo, raggiungendo un picco di periodo tardo fasi di proliferazione endometriale. Dopo l'ovulazione, si verifica una rapida diminuzione della concentrazione dei recettori nucleari dell'estradiolo, continuando fino alla fase secretoria tardiva, quando la loro espressione diventa significativamente inferiore rispetto all'inizio del ciclo. È stato stabilito che l'induzione della formazione di recettori sia per l'estradiolo che per il progesterone dipende dalla concentrazione di estradiolo nei tessuti. presto fase proliferativa il contenuto di recettori per il progesterone è inferiore a quello per l'estradiolo, ma poi si verifica un aumento preovulatoria del livello dei recettori del progesterone.
Dopo l'ovulazione, il livello dei recettori nucleari per il progesterone raggiunge il massimo per l'intero ciclo. Nella fase proliferativa, l'estradiolo stimola direttamente la formazione dei recettori del progesterone, il che spiega la mancanza di relazione tra i livelli plasmatici di progesterone e il contenuto dei suoi recettori nell'endometrio. La regolazione della concentrazione locale di estradiolo e progesterone è mediata in larga misura dalla comparsa di vari enzimi durante il ciclo mestruale. Il contenuto di estrogeni nell'endometrio dipende non solo dal loro livello nel sangue, ma anche dall'istruzione. L'endometrio di una donna è in grado di sintetizzare gli estrogeni convertendo androstenedione e testosterone con la partecipazione dell'aromatasi (aromatizzazione). Questa fonte locale di estrogeni migliora l'estrogenizzazione delle cellule endometriali che caratterizza la fase proliferativa. Durante questa fase si nota la più alta aromatizzazione degli androgeni e la più bassa attività degli enzimi che metabolizzano gli estrogeni. Recentemente è stato accertato che l'endometrio è in grado di secernere prolattina, che è del tutto identica all'ipofisi. La sintesi della prolattina da parte dell'endometrio inizia nella seconda metà della fase luteale (attivata dal progesterone) e coincide con la decidualizzazione delle cellule stromali. L'attività ciclica del sistema riproduttivo è determinata dai principi di diretto e feedback, forniti da specifici recettori ormonali in ciascuno dei collegamenti. Un collegamento diretto è l'effetto stimolante dell'ipotalamo sulla ghiandola pituitaria e la successiva formazione di steroidi sessuali nell'ovaio. Il feedback è determinato dall'influenza della maggiore concentrazione di steroidi sessuali sui livelli sovrastanti. Nell'interazione dei collegamenti del sistema riproduttivo si distinguono i loop "lunghi", "brevi" e "ultracorti". L'ansa "lunga" è l'effetto attraverso i recettori del sistema ipotalamo-ipofisario sulla produzione di ormoni sessuali. L'ansa "corta" definisce la connessione tra la ghiandola pituitaria e l'ipotalamo. Anello "ultra corto" - connessione tra l'ipotalamo e cellule nervose, che effettuano la regolazione locale con l'ausilio di neurotrasmettitori, neuropeptidi, neuromodulatori e stimoli elettrici.
Valutazione dello stato del sistema riproduttivo secondo test di diagnostica funzionale. Per molti anni nella pratica ginecologica sono stati utilizzati i cosiddetti test di diagnostica funzionale dello stato del sistema riproduttivo. Il valore di questi studi piuttosto semplici è stato preservato fino ai giorni nostri. Le più comunemente utilizzate sono la misurazione della temperatura basale, la valutazione del fenomeno "pupilla" e del muco cervicale (cristallizzazione, distensibilità), nonché il calcolo dell'indice cariopicnotico (KPI, %) dell'epitelio vaginale.
Test della temperatura basale si basa sulla capacità del progesterone (in maggiore concentrazione) di provocare una ristrutturazione del lavoro del centro di termoregolazione ipotalamica, che porta a una reazione ipertermica transitoria. La temperatura viene misurata quotidianamente nel retto al mattino senza alzarsi dal letto. I risultati vengono visualizzati graficamente. Con un normale ciclo mestruale a due fasi, la temperatura basale nella fase del progesterone aumenta di 0,4--0,8 ° C. Il giorno delle mestruazioni o 1 giorno prima dell'inizio, la temperatura basale diminuisce. Un ciclo persistente a due fasi (la temperatura basale deve essere misurata in 2-3 cicli mestruali) indica che si è verificata l'ovulazione e un corpo luteo funzionalmente attivo. L'assenza di un aumento della temperatura nella seconda fase del ciclo indica l'anovulazione, un ritardo nell'aumento e / o la sua breve durata (un aumento della temperatura di 2--7 giorni) - un accorciamento della fase luteale, un aumento insufficiente (di 0,2-0,3 ° C) - - per insufficienza della funzione del corpo luteo. Un risultato falso positivo (un aumento della temperatura basale in assenza di un corpo luteo) può essere in acuto e infezioni croniche, con alcuni cambiamenti nel sistema nervoso centrale, accompagnati da una maggiore eccitabilità. Sintomo "pupilla" riflette la quantità e le condizioni della secrezione mucosa nel canale cervicale, che dipendono dalla saturazione di estrogeni del corpo. Il numero più grande muco cervicale formato durante l'ovulazione, il più piccolo - prima delle mestruazioni. Il fenomeno "pupilla" si basa sull'espansione dell'os esterno del canale cervicale dovuto all'accumulo di muco vitreo trasparente nella cervice. Nei giorni preovulatori, l'apertura esterna dilatata del canale cervicale ricorda una pupilla. Il fenomeno "pupilla", a seconda della sua gravità, è stimato in 1-3 plus. Il test non può essere utilizzato alterazioni patologiche cervice.
Valutazione della qualità del muco cervicale riflette la sua cristallizzazione e il grado di tensione. La cristallizzazione (il fenomeno della "felce") del muco cervicale durante l'essiccazione è più pronunciata durante l'ovulazione, quindi diminuisce gradualmente ed è completamente assente prima delle mestruazioni. In punti (da 1 a 3) viene valutata anche la cristallizzazione del muco essiccato all'aria. La tensione del muco cervicale dipende dalla saturazione di estrogeni. Korntsang da canale cervicale il muco viene rimosso, le mascelle dello strumento vengono allontanate, determinando il grado di tensione. Prima delle mestruazioni, la lunghezza del filo è massima (12 cm). Il muco può essere influenzato negativamente dai processi infiammatori negli organi genitali e dagli squilibri ormonali.
Indice cariopicotico. Le fluttuazioni cicliche degli ormoni ovarici sono associate a cambiamenti composizione cellulare mucosa dell'endometrio. In uno striscio dalla vagina, in base alle caratteristiche morfologiche, si distinguono 4 tipi di cellule epiteliali stratificate squamose:
- a) cheratinizzante;
- b) intermedio;
- c) parabasale;
- d) basale. L'indice karyopyknotic (KPI) è il rapporto tra il numero di cellule con un nucleo picnotico (cioè cellule cheratinizzanti) e il numero totale di cellule epiteliali in uno striscio, espresso in percentuale.
Nella fase follicolare del ciclo mestruale, l'IPC è del 20–40%, nei giorni preovulatori sale all'80–88% e nella fase luteale del ciclo diminuisce al 20–25%. Pertanto, i rapporti quantitativi degli elementi cellulari negli strisci della mucosa vaginale consentono di giudicare la saturazione del corpo con gli estrogeni.
Elenco delle abbreviazioni:
ADH - ormone antidiuretico
ACTH - corticoliberina
aRG-GN - agonista dell'ormone di rilascio delle gonadotropine
LH - ormone luteinizzante
OP - ossiprogesterone
RG-GN - ormone di rilascio delle gonadotropine
STH - somatoliberina
VEGF - fattore di crescita dell'endotelio vascolare
TSH - ormone tireotropo (tiroliberina)
FSH - ormone follicolo-stimolante
FGF - fattore di crescita fibroplastico
Ciclo mestruale normale
Mestruazioni- questa è una scarica sanguinolenta dal tratto genitale di una donna, che si verifica periodicamente a seguito del rigetto dello strato funzionale dell'endometrio alla fine di un ciclo mestruale in due fasi.
Il complesso dei processi ciclici che si verificano nel corpo femminile e si manifestano esternamente dalle mestruazioni è chiamato ciclo mestruale. Le mestruazioni iniziano come risposta a un cambiamento nel livello di steroidi prodotti dalle ovaie.
Segni clinici di un normale ciclo mestrualeLa durata del ciclo mestruale nel periodo riproduttivo attivo di una donna è in media di 28 giorni. Una durata del ciclo da 21 a 35 giorni è considerata normale. Si osservano ampi intervalli durante la pubertà e la menopausa, che possono essere una manifestazione di anovulazione, che può verificarsi più spesso in questo momento.
Di solito le mestruazioni durano dai 3 ai 7 giorni, la quantità di sangue persa è trascurabile. Accorciamento o allungamento del sanguinamento mestruale, così come l'aspetto di scarso o mestruazioni abbondanti può servire come manifestazione di una serie di malattie ginecologiche.
Caratteristiche di un normale ciclo mestruale:
Durata: 28±7 giorni;
Durata del sanguinamento mestruale: 4±2 giorni;
Volume di perdita di sangue durante le mestruazioni: 20-60 ml * ;
Perdita media di ferro: 16 mg
*
Il 95% delle donne sane perde meno di 60 ml di sangue con ogni mestruazione. La perdita di sangue di oltre 60-80 ml è combinata con una diminuzione dell'emoglobina, dell'ematocrito e del ferro sierico.
Fisiologia del sanguinamento mestruale:
Immediatamente prima delle mestruazioni, si sviluppa uno spasmo pronunciato delle arteriole a spirale. Dopo la dilatazione delle arteriole a spirale, inizia il sanguinamento mestruale. Inizialmente, l'adesione piastrinica nei vasi endometriali viene soppressa, ma poi, con il progredire della trasudazione del sangue, le estremità danneggiate dei vasi vengono sigillate con trombi intravascolari, costituiti da piastrine e fibrina. 20 ore dopo l'inizio delle mestruazioni, quando la maggior parte dell'endometrio è già stata strappata via, si sviluppa uno spasmo pronunciato delle arteriole a spirale, grazie al quale si ottiene l'emostasi. La rigenerazione dell'endometrio inizia 36 ore dopo l'inizio delle mestruazioni, nonostante il rigetto dell'endometrio non sia ancora completamente completato.
La regolazione del ciclo mestruale è un complesso meccanismo neuroumorale, che viene effettuato con la partecipazione di 5 principali anelli di regolazione. Questi includono: la corteccia cerebrale, i centri sottocorticali (ipotalamo), la ghiandola pituitaria, le ghiandole sessuali, gli organi e i tessuti periferici (utero, tube di Falloppio, vagina, ghiandole mammarie, follicoli piliferi, ossa, tessuto adiposo). Questi ultimi sono detti organi bersaglio, per la presenza di recettori sensibili all'azione degli ormoni che l'ovaio produce durante il ciclo mestruale. Recettori del citosol - recettori del citoplasma, hanno una stretta specificità per estradiolo, progesterone, testosterone, mentre i recettori nucleari possono essere accettori di molecole come insulina, glucagone, aminopeptidi.
I recettori per gli ormoni sessuali si trovano in tutte le strutture del sistema riproduttivo, così come nel sistema nervoso centrale, nella pelle, nel tessuto adiposo e osseo e nella ghiandola mammaria. Una molecola di ormone steroideo libero viene catturata da uno specifico recettore del citosol di natura proteica, il complesso risultante viene traslocato nel nucleo cellulare. Nel nucleo appare un nuovo complesso con un recettore di proteine nucleari; questo complesso si lega alla cromatina, che regola la trascrizione dell'mRNA ed è coinvolta nella sintesi di una specifica proteina tissutale. Il mediatore intracellulare - acido adenosina monofosforico ciclico (cAMP) regola il metabolismo nelle cellule del tessuto bersaglio in base alle esigenze dell'organismo in risposta agli effetti degli ormoni. La maggior parte degli ormoni steroidei (circa l'80% è nel sangue e viene trasportato in una forma legata. Il loro trasporto è effettuato da proteine speciali: globuline leganti gli steroidi e sistemi di trasporto non specifici (albumine ed eritrociti). In una forma legata , gli steroidi sono inattivi, pertanto globuline, albumine ed eritrociti possono essere considerati una sorta di sistema tampone che controlla l'accesso degli steroidi ai recettori delle cellule bersaglio.
I cambiamenti funzionali ciclici che si verificano nel corpo di una donna possono essere suddivisi condizionatamente in cambiamenti nel sistema ipotalamo-ipofisi-ovaie (ciclo ovarico) e nell'utero, principalmente nella sua membrana mucosa (ciclo uterino).
Insieme a questo, di regola, si verificano spostamenti ciclici in tutti gli organi e sistemi di una donna, in particolare nel sistema nervoso centrale, nel sistema cardiovascolare, nel sistema di termoregolazione, processi metabolici eccetera.
Ipotalamo
L'ipotalamo è la parte del cervello situata sopra il chiasma ottico e che forma la parte inferiore del terzo ventricolo. È un componente vecchio e stabile del sistema nervoso centrale, organizzazione generale che è cambiato poco nel corso dell'evoluzione umana. Strutturalmente e funzionalmente, l'ipotalamo è correlato alla ghiandola pituitaria. Ci sono tre regioni ipotalamiche: anteriore, posteriore e intermedia. Ogni area è formata da nuclei: accumuli di corpi di neuroni di un certo tipo.
Oltre alla ghiandola pituitaria, l'ipotalamo colpisce il sistema limbico (amigdala, ippocampo), il talamo e il ponte. Questi dipartimenti influenzano anche direttamente o indirettamente l'ipotalamo.
L'ipotalamo secerne liberine e statine. Questo processo è regolato da ormoni che chiudono tre circuiti di feedback: lungo, corto e ultracorto. Un lungo ciclo di feedback è fornito dagli ormoni sessuali circolanti che si legano ai corrispondenti recettori nell'ipotalamo, uno corto: gli ormoni dell'adenoipofisi, uno ultracorto: liberine e statine. Liberine e statine regolano l'attività dell'adenoipofisi. La gonadoliberina stimola la secrezione di LH e FSH, corticoliberina - ACTH, somatoliberina (STG), tiroliberina (TSH). Oltre alle liberine e alle statine, nell'ipotalamo vengono sintetizzati l'ormone antidiuretico e l'ossitocina. Questi ormoni vengono trasportati nella neuroipofisi, da dove entrano nel flusso sanguigno.
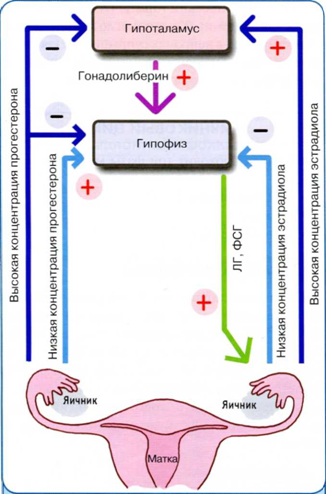
A differenza dei capillari di altre aree del cervello, i capillari dell'imbuto dell'ipotalamo sono fenestrati. Costituiscono la rete capillare primaria del sistema portale.
Negli anni 70-80. una serie di studi sperimentali è stata condotta sulle scimmie, che ha permesso di identificare differenze nella funzione delle strutture neurosecretorie dell'ipotalamo di primati e roditori. Nei primati e nell'uomo, i nuclei arcuati dell'ipotalamo mediobasale sono l'unico sito per la formazione e il rilascio di RG-LH, che è responsabile della funzione gonadotropica della ghiandola pituitaria. La secrezione di RG-LH è programmata geneticamente e avviene con un certo ritmo pulsante con una frequenza di circa una volta all'ora. Questo ritmo è chiamato circolare (ora-esima). La regione dei nuclei arcuati dell'ipotalamo è chiamata oscillatore arcuato. La natura circolare della secrezione di RG-LH è stata confermata dalla determinazione diretta di essa nel sangue del sistema portale del peduncolo ipofisario e della vena giugulare nelle scimmie e nel sangue delle donne con ciclo ovulatorio.
Ormoni dell'ipotalamo
L'ormone di rilascio LH è stato isolato, sintetizzato e descritto in dettaglio. Ad oggi, non è stato possibile isolare e sintetizzare la folliberina. L'RG-LH e i suoi analoghi sintetici hanno la capacità di stimolare il rilascio di LH e FSH dalla ghiandola pituitaria anteriore, pertanto è attualmente accettato un termine per i liberins gonadotropici ipotalamici: l'ormone di rilascio delle gonadotropine (RG-GN).
La gonadoliberina stimola la secrezione di FSH e LH. È un decapeptide secreto dai neuroni del nucleo dell'infundibolo. La gonadoliberina viene secreta non costantemente, ma in modalità pulsata. Viene distrutto molto rapidamente dalle proteasi (l'emivita è di 2-4 minuti), quindi la sua impulso deve essere regolare. La frequenza e l'ampiezza delle emissioni di GnRH cambiano durante il ciclo mestruale. La fase follicolare è caratterizzata da frequenti fluttuazioni della piccola ampiezza del livello di gonadoliberina nel siero del sangue. Verso la fine della fase follicolare, la frequenza e l'ampiezza delle oscillazioni aumentano, per poi diminuire durante la fase luteale.
pituitario
Ci sono due lobi nella ghiandola pituitaria: anteriore - adenoipofisi e posteriore - neuroipofisi. La neuroipofisi è di origine neurogena e rappresenta una continuazione dell'imbuto dell'ipotalamo. La neuroipofisi riceve il suo apporto di sangue dalle arterie pituitarie inferiori. L'adenoipofisi si sviluppa dall'ectoderma della sacca di Rathke, quindi è costituita da un epitelio ghiandolare e non ha connessione diretta con l'ipotalamo. Sintetizzate nell'ipotalamo, le liberine e le statine entrano nell'adenoipofisi attraverso uno speciale sistema di cancello. È la principale fonte di afflusso di sangue all'adenoipofisi. Il sangue entra nel sistema portale principalmente attraverso le arterie pituitarie superiori. Nella regione dell'imbuto dell'ipotalamo, formano la rete capillare primaria del sistema portale, da cui si formano le vene porta, che entrano nell'adenoipofisi e danno origine a una rete capillare secondaria. È possibile il flusso inverso di sangue attraverso il sistema portale. Le caratteristiche dell'afflusso di sangue e l'assenza della barriera ematoencefalica nell'imbuto dell'ipotalamo forniscono una connessione bidirezionale tra l'ipotalamo e la ghiandola pituitaria. A seconda della colorazione con ematossilina ed eosina, le cellule secretorie dell'adenoipofisi sono divise in cromofile (acidofile) e basofile (cromofobiche). Le cellule acidofile secernono l'ormone della crescita e la prolattina, cellule basofile - FSH, LH, TSH, ACTH
ormoni ipofisari
L'adenoipofisi produce GH, prolattina, FSH, LH, TSH e ACTH. FSH e LH regolano la secrezione degli ormoni sessuali, TSH - la secrezione degli ormoni tiroidei, ACTH - la secrezione degli ormoni della corteccia surrenale. STH stimola la crescita, ha un effetto anabolico. La prolattina stimola la crescita delle ghiandole mammarie durante la gravidanza e l'allattamento dopo il parto.
LH e FSH sono sintetizzati dalle cellule gonadotropiche dell'adenoipofisi e svolgono un ruolo importante nello sviluppo dei follicoli ovarici. Strutturalmente, sono classificati come glicoproteine. L'FSH stimola la crescita del follicolo, la proliferazione delle cellule della granulosa, induce la formazione di recettori LH sulla superficie delle cellule della granulosa. Sotto l'influenza dell'FSH, il contenuto di aromatasi nel follicolo in maturazione aumenta. L'LH stimola la formazione di androgeni (precursori degli estrogeni) nelle cellule della teca, insieme all'FSH favorisce l'ovulazione e stimola la sintesi del progesterone nelle cellule della granulosa luteinizzata del follicolo ovulato.
La secrezione di LH e FSH è variabile e modulata dagli ormoni ovarici, in particolare estrogeni e progesterone.
In questo modo, basso livello gli estrogeni hanno un effetto soppressivo sull'LH, mentre uno alto stimola la sua produzione da parte della ghiandola pituitaria. Nella fase follicolare tardiva, i livelli sierici di estrogeni sono piuttosto elevati, l'effetto di feedback positivo è triplicato, il che contribuisce alla formazione di un picco di LH preovulatoria. E, al contrario, durante la terapia con contraccettivi combinati, il livello di estrogeni nel siero del sangue rientra nei limiti che determinano un feedback negativo, che porta ad una diminuzione del contenuto di gonadotropine.
Il meccanismo di feedback positivo porta ad un aumento della concentrazione e della produzione di RG-GN nei recettori.
Contrariamente all'effetto degli estrogeni, bassi livelli di progesterone hanno un feedback positivo sulla secrezione di LH e FSH da parte della ghiandola pituitaria. Queste condizioni esistono appena prima dell'ovulazione e portano al rilascio di FSH. L'alto livello di progesterone, che si nota nella fase luteale, riduce la produzione ipofisaria di gonadotropine. Una piccola quantità di progesterone stimola il rilascio di gonadotropine a livello della ghiandola pituitaria. L'effetto di feedback negativo del progesterone si manifesta con una diminuzione della produzione di RG-GN e una diminuzione della sensibilità a RG-GN a livello della ghiandola pituitaria. L'effetto di feedback positivo del progesterone si verifica sulla ghiandola pituitaria e include una maggiore sensibilità all'RH-GN. Gli estrogeni e il progesterone non sono gli unici ormoni che influenzano la secrezione di gonadotropine da parte della ghiandola pituitaria. Gli ormoni inibina e attivina hanno lo stesso effetto. L'inibina sopprime la secrezione ipofisaria di FSH, mentre l'attivina la stimola.
Prolattinaè un polipeptide costituito da 198 residui di amminoacidi, sintetizzati dalle cellule lattotropiche dell'adenoipofisi. La secrezione di prolattina è controllata dalla dopamina. È sintetizzato nell'ipotalamo e inibisce la secrezione di prolattina. La prolattina ha una varietà di effetti sul corpo di una donna. Il suo principale ruolo biologico è la crescita delle ghiandole mammarie e la regolazione dell'allattamento. Ha anche un effetto mobilizzante del grasso e ha un effetto ipotensivo. Un aumento della secrezione di prolattina è una delle cause più comuni di infertilità, poiché un aumento del suo livello nel sangue inibisce la steroidogenesi nelle ovaie e lo sviluppo dei follicoli.
Ossitocina- un peptide costituito da 9 residui di amminoacidi. Si forma nei neuroni della grande parte cellulare dei nuclei paraventricolari dell'ipotalamo. I principali obiettivi dell'ossitocina nell'uomo sono le fibre muscolari lisce dell'utero e le cellule mioepiteliali delle ghiandole mammarie.
Ormone antidiuretico(ADH) è un peptide costituito da 9 residui di amminoacidi. Sintetizzato nei neuroni del nucleo sovraottico dell'ipotalamo. La funzione principale dell'ADH è la regolazione del BCC, della pressione sanguigna e dell'osmolalità plasmatica.
Ciclo ovarico
Le ovaie attraversano tre fasi del ciclo mestruale:
- fase follicolare;
- ovulazione;
- fase luteale.
Fase follicolare:
Uno dei punti salienti della fase follicolare del ciclo mestruale è lo sviluppo dell'uovo. L'ovaio femminile è organo complesso, costituito da molti componenti, a seguito dell'interazione di cui vengono secreti gli ormoni steroidei sessuali e si forma un uovo pronto per la fecondazione in risposta alla secrezione ciclica delle gonadotropine.
Steroidogenesi
L'attività ormonale dal follicolo preantrale a quello periovulatorio è stata descritta come la teoria delle "due cellule, due gonadotropine". La steroidogenesi si verifica in due cellule del follicolo: le cellule della teca e della granulosa. Nelle cellule della teca, LH stimola la produzione di androgeni dal colesterolo. Nelle cellule della granulosa, l'FSH stimola la conversione degli androgeni risultanti in estrogeni (aromatizzazione). Oltre all'effetto aromatizzante, l'FSH è anche responsabile della proliferazione delle cellule della granulosa. Sebbene siano noti altri mediatori nello sviluppo dei follicoli ovarici, questa teoria è la principale per comprendere i processi che si verificano nel follicolo ovarico. È stato rivelato che entrambi gli ormoni sono necessari per un ciclo normale con un livello sufficiente di estrogeni.
La produzione di androgeni nei follicoli può anche regolare lo sviluppo del follicolo preantrale. Un basso livello di androgeni migliora il processo di aromatizzazione, quindi aumenta la produzione di estrogeni e viceversa, un livello alto inibisce il processo di aromatizzazione e provoca atresia del follicolo. Un equilibrio di FSH e LH è essenziale per sviluppo iniziale follicolo. La condizione ottimale per stato iniziale lo sviluppo del follicolo è basso LH e alto FSH, che si verifica all'inizio del ciclo mestruale. Se il livello di LH è alto, le cellule della teca producono grandi quantità di androgeni, causando atresia follicolare.
Selezione del follicolo dominante
La crescita del follicolo è accompagnata dalla secrezione di ormoni steroidei sessuali sotto l'influenza di LH e FSH. Queste gonadotropine proteggono il gruppo del follicolo preantrale dall'atresia. Tuttavia, normalmente solo uno di questi follicoli si sviluppa nel follicolo preovulatorio, che viene quindi rilasciato e diventa dominante.
Il follicolo dominante nella fase follicolare media è il più grande e sviluppato nell'ovaio. Già nei primi giorni del ciclo mestruale ha un diametro di 2 mm e entro 14 giorni dal momento dell'ovulazione aumenta a una media di 21 mm. Durante questo periodo, si verifica un aumento di 100 volte del volume del fluido follicolare, il numero di cellule della granulosa che rivestono la membrana basale aumenta da 0,5x10 6 a 50x10 6 . Questo follicolo ha la più alta attività aromatizzante e la più alta concentrazione di recettori LH indotti dall'FSH, quindi il follicolo dominante secerne le quantità più elevate di estradiolo e inibina. Inoltre, l'inibina migliora la sintesi degli androgeni sotto l'influenza dell'LH, che è un substrato per la sintesi dell'estradiolo.
A differenza del livello di FSH, che diminuisce all'aumentare della concentrazione di estradiolo, il livello di LH continua ad aumentare (a basse concentrazioni, l'estradiolo inibisce la secrezione di LH). È la stimolazione a lungo termine degli estrogeni che prepara il picco ovulatoria di LH. Allo stesso tempo, il follicolo dominante si prepara all'ovulazione: sotto l'azione locale degli estrogeni e dell'FSH, aumenta il numero di recettori LH sulle cellule della granulosa. Il rilascio di LH porta all'ovulazione, alla formazione di un corpo luteo e ad un aumento della secrezione di progesterone. L'ovulazione avviene 10-12 ore dopo il picco di LH o 32-35 ore dopo l'inizio dell'aumento del suo livello. Di solito un solo follicolo ovula.
Durante la selezione del follicolo, i livelli di FSH diminuiscono in risposta a effetto negativo dagli estrogeni, quindi il follicolo dominante è l'unico che continua a svilupparsi con livelli di FSH in calo
La connessione ovarico-ipofisi è determinante nella scelta del follicolo dominante e nello sviluppo dell'atresia dei follicoli rimanenti.
inibina e attivina
La crescita e lo sviluppo dell'uovo, il funzionamento del corpo luteo avviene attraverso l'interazione di meccanismi autocrini e paracrini. Va notato due ormoni follicolari che svolgono un ruolo significativo nella steroidogenesi: l'inibina e l'attivina.
L'inibina è un ormone peptidico prodotto dalle cellule della granulosa dei follicoli in crescita che riduce la produzione di FSH. Inoltre, influisce sulla sintesi degli androgeni nell'ovaio. L'inibina agisce sulla follicologenesi nel modo seguente: riducendo l'FSH a un livello in cui si sviluppa solo un follicolo dominante.
L'attivina è un ormone peptidico prodotto nelle cellule della granulosa dei follicoli e della ghiandola pituitaria. Secondo alcuni autori, l'attivina è prodotta anche dalla placenta. L'attivina aumenta la produzione di FSH da parte della ghiandola pituitaria, migliora il legame di FSH alle cellule della granulosa.
Fattori di crescita insulino-simili
I fattori di crescita insulino-simili (IGF-1 e IGF-2) sono sintetizzati nel fegato sotto l'influenza dell'ormone della crescita e, possibilmente, nelle cellule della granulosa dei follicoli, agiscono come regolatori paracrini. Prima dell'ovulazione, il contenuto di IGF-1 e IGF-2 nel liquido follicolare aumenta a causa dell'aumento della quantità di liquido stesso nel follicolo dominante. L'IGF-1 è coinvolto nella sintesi dell'estradiolo. L'IGF-2 (epidermico) inibisce la sintesi degli steroidi nelle ovaie.
Ovulazione:
Il picco ovulatoria di LH porta ad un aumento della concentrazione di prostaglandine e dell'attività proteasica nel follicolo. Il processo di ovulazione stesso è una rottura della membrana basale del follicolo dominante e sanguinamento dai capillari distrutti che circondano le cellule della teca. I cambiamenti nella parete del follicolo preovulatorio, che ne assicurano l'assottigliamento e la rottura, si verificano sotto l'influenza dell'enzima collagenasi; un certo ruolo è svolto anche dalle prostaglandine contenute nel liquido follicolare, dagli enzimi proteolitici formatisi nelle cellule della granulosa, dall'ossitopina e dalla relaxina. Di conseguenza, si forma un piccolo foro nella parete del follicolo, attraverso il quale l'uovo viene rilasciato lentamente. Le misurazioni dirette hanno dimostrato che la pressione all'interno del follicolo non aumenta durante l'ovulazione.
Al termine della fase follicolare, l'FSH agisce sui recettori LH nelle cellule della granulosa. Gli estrogeni sono un cofattore obbligatorio in questo effetto. Quando il follicolo dominante si sviluppa, la produzione di estrogeni aumenta. Di conseguenza, la produzione di estrogeni è sufficiente per ottenere la secrezione di LH da parte della ghiandola pituitaria, che porta ad un aumento del suo livello. L'aumento avviene dapprima molto lentamente (dall'8° al 12° giorno del ciclo), poi rapidamente (dopo il 12° giorno del ciclo). Durante questo periodo, LH attiva la luteinizzazione delle cellule della granulosa nel follicolo dominante. Pertanto, il progesterone viene rilasciato. Inoltre, il progesterone migliora l'effetto degli estrogeni sulla secrezione di LH ipofisario, portando ad un aumento del suo livello.
L'ovulazione avviene entro 36 ore dall'inizio del picco di LH. La definizione di un picco di LH è una di migliori pratiche, che determina l'ovulazione e viene eseguita utilizzando il dispositivo "determinante dell'ovulazione".
Il picco periovulatorio dell'FSH si verifica probabilmente come risultato dell'effetto positivo del progesterone. Oltre all'aumento di LH, FSH ed estrogeni, c'è anche un aumento dei livelli sierici di androgeni durante l'ovulazione. Questi androgeni vengono rilasciati come risultato dell'effetto stimolatore dell'LH sulle cellule della teca, specialmente nel follicolo non dominante.
L'aumento degli androgeni ha un effetto sull'aumento della libido, confermando che questo periodo è il più fertile nelle donne.
I livelli di LH stimolano la meiosi dopo che lo sperma è entrato nell'uovo. Quando un ovocita viene rilasciato dall'ovaio durante l'ovulazione, la parete del follicolo viene distrutta. Questo è regolato da LH, FSH e progesterone, che stimolano l'attività degli enzimi proteolitici come gli attivatori del plasminogeno (che rilasciano plasmina, che stimola l'attività della collagenasi) e le prostaglandine. Le prostaglandine non solo aumentano l'attività degli enzimi proteolitici, ma contribuiscono anche alla comparsa di una reazione infiammatoria nella parete follicolare e stimolano l'attività della muscolatura liscia, che contribuisce al rilascio dell'ovocita.
L'importanza delle prostaglandine nel processo di ovulazione è stata dimostrata da studi che indicano che una diminuzione del rilascio di prostaglandine può portare a un ritardo nel rilascio dell'ovocita dall'ovaio durante la normale steroidogenesi (sindrome del follicolo luteinizzato non in via di sviluppo - SNLF). Poiché il SNLF è spesso la causa dell'infertilità, si consiglia alle donne che desiderano una gravidanza di evitare l'assunzione di inibitori delle prostaglandine sintetizzate.
fase luteale:
La struttura del corpo luteo
Dopo il rilascio dell'uovo dall'ovaio, i capillari in formazione crescono rapidamente nella cavità del follicolo; le cellule della granulosa subiscono la luteinizzazione: un aumento del citoplasma in esse e la formazione di inclusioni lipidiche. Le cellule della granulosa e i tecociti formano il corpo luteo, il principale regolatore della fase luteale del ciclo mestruale. Le cellule che formavano la parete del follicolo accumulano lipidi e il pigmento giallo luteina e iniziano a secernere progesterone, estradiolo-2 e inibina. Una potente rete vascolare contribuisce all'ingresso degli ormoni del corpo luteo nella circolazione sistemica. Un vero e proprio corpo luteo si sviluppa solo quando nel follicolo preovulatore si forma un numero adeguato di cellule della granulosa con un alto contenuto di recettori LH. L'aumento delle dimensioni del corpo luteo dopo l'ovulazione si verifica principalmente a causa dell'aumento delle dimensioni delle cellule della granulosa, mentre il loro numero non aumenta per l'assenza di mitosi. Nell'uomo, il corpo luteo secerne non solo progesterone, ma anche estradiolo e androgeni. I meccanismi di regressione del corpo luteo non sono ben compresi. È noto che le prostaglandine hanno un effetto luteolitico.

Riso. Immagine ecografica del corpo luteo "in fiore" durante la gravidanza 6 settimane. 4 giorni. Modalità mappatura energetica.
Regolazione ormonale della fase luteale
Se la gravidanza non si verifica, si verifica l'involuzione del corpo luteo. Questo processo è regolato da un meccanismo di feedback negativo: gli ormoni (progesterone ed estradiolo) secreti dal corpo luteo agiscono sulle cellule gonadotropiche della ghiandola pituitaria, sopprimendo la secrezione di FSH e LH. L'inibina inibisce anche la secrezione di FSH. La diminuzione dei livelli di FSH, così come l'azione locale del progesterone, impedisce lo sviluppo di un gruppo di follicoli primordiali.
L'esistenza del corpo luteo dipende dal livello di secrezione di LH. Quando diminuisce, di solito 12-16 giorni dopo l'ovulazione, si verifica l'involuzione del corpo luteo. Al suo posto si forma un corpo bianco. Il meccanismo dell'involuzione è sconosciuto. Molto probabilmente, è dovuto a influenze paracrine. Quando il corpo luteo si evolve, i livelli di estrogeni e progesterone diminuiscono, portando a una maggiore secrezione di ormoni gonadotropici. Quando il contenuto di FSH e LH aumenta, inizia a svilupparsi un nuovo gruppo di follicoli.
Se è avvenuta la fecondazione, l'esistenza del corpo luteo e la secrezione di progesterone è supportata dalla gonadotropina corionica. Pertanto, l'impianto dell'embrione porta a cambiamenti ormonali che preservano il corpo luteo.
La durata della fase luteale nella maggior parte delle donne è costante ed è di circa 14 giorni.
Ormoni ovarici
Il complesso processo di biosintesi degli steroidi termina con la formazione di estradiolo, testosterone e progesterone. I tessuti delle ovaie che producono steroidi sono cellule della granulosa che rivestono la cavità del follicolo, cellule della teca interna e, in misura molto minore, lo stroma. Le cellule della granulosa e le cellule della teca sono sinergicamente coinvolte nella sintesi degli estrogeni, le cellule della membrana tecale sono la principale fonte di androgeni, che si formano anche in piccole quantità nello stroma; il progesterone è sintetizzato nelle cellule della teca e nelle cellule della granulosa.
Nell'ovaio vengono secreti 60-100 mcg di estradiolo (E2) nella prima fase follicolare del ciclo mestruale, 270 mcg nella fase luteale e 400-900 mcg al giorno al momento dell'ovulazione. Circa il 10% di E2 è aromatizzato nell'ovaio dal testosterone. La quantità di estrone formata nella fase follicolare iniziale è di 60-100 mcg, al momento dell'ovulazione la sua sintesi aumenta a 600 mcg al giorno. Solo la metà della quantità di estrone viene prodotta nell'ovaio. L'altra metà è aromatizzata a E2. L'estriolo è un metabolita inattivo dell'estradiolo e dell'estrone.
Il progesterone viene prodotto nell'ovaio a 2 mg/die durante la fase follicolare e 25 mg/die durante la fase luteale del ciclo mestruale. Nel processo di metabolismo, il progesterone nell'ovaio si trasforma in 20-deidroprogesterone, che ha un'attività biologica relativamente bassa.
Nell'ovaio vengono sintetizzati i seguenti androgeni: androstenedione (un precursore del testosterone) in una quantità di 1,5 mg/die (la stessa quantità di androstenedione si forma nelle ghiandole surrenali). Circa 0,15 mg di testosterone si formano dall'androstenedione, approssimativamente la stessa quantità si forma nelle ghiandole surrenali.
Una breve panoramica dei processi che si verificano nelle ovaie
Fase follicolare:
LH stimola la produzione di androgeni nelle cellule della teca.
L'FSH stimola la produzione di estrogeni nelle cellule della granulosa.
Il follicolo più sviluppato nel mezzo della fase follicolare diventa dominante.
L'aumento della produzione di estrogeni e inibina nel follicolo dominante sopprime il rilascio di FSH da parte della ghiandola pituitaria.
Una diminuzione dei livelli di FSH provoca atresia di tutti i follicoli tranne quello dominante.
Ovulazione:
L'FSH induce i recettori LH.
Gli enzimi proteolitici nel follicolo portano alla distruzione della sua parete e al rilascio dell'ovocita.
fase luteale:
Il corpo luteo è formato da cellule della granulosa e della teca conservate dopo l'ovulazione.
Il progesterone, secreto dal corpo luteo, è l'ormone dominante. In assenza di gravidanza, la luteolisi si verifica 14 giorni dopo l'ovulazione.
ciclo uterino
L'endometrio è costituito da due strati: funzionale e basale. Lo strato funzionale cambia struttura sotto l'azione degli ormoni sessuali e, se la gravidanza non si verifica, viene rifiutato durante le mestruazioni.
Fase proliferativa:
L'inizio del ciclo mestruale è considerato il 1° giorno delle mestruazioni. Alla fine delle mestruazioni, lo spessore dell'endometrio è di 1-2 mm. L'endometrio è costituito quasi esclusivamente dallo strato basale. Le ghiandole sono strette, diritte e corte, rivestite da un epitelio cilindrico basso, il citoplasma delle cellule stromali è quasi lo stesso. All'aumentare del livello di estradiolo, si forma uno strato funzionale: l'endometrio si prepara all'impianto dell'embrione. Le ghiandole si allungano e diventano tortuose. Il numero di mitosi aumenta. Con la proliferazione, l'altezza delle cellule epiteliali aumenta e l'epitelio stesso da una singola fila diventa a più file al momento dell'ovulazione. Lo stroma è edematoso e allentato, i nuclei delle cellule e il volume del citoplasma aumentano in esso. Le navi sono moderatamente tortuose.
fase secretoria:
Normalmente, l'ovulazione avviene il 14° giorno del ciclo mestruale. La fase secretoria è caratterizzata da alti livelli di estrogeni e progesterone. Tuttavia, dopo l'ovulazione, il numero di recettori degli estrogeni nelle cellule endometriali diminuisce. La proliferazione dell'endometrio viene gradualmente inibita, la sintesi del DNA diminuisce e il numero di mitosi diminuisce. Pertanto, il progesterone ha un effetto predominante sull'endometrio nella fase secretoria.
I vacuoli contenenti glicogeno compaiono nelle ghiandole dell'endometrio, che vengono rilevati utilizzando la reazione PAS. Al 16° giorno del ciclo, questi vacuoli sono abbastanza grandi, presenti in tutte le cellule e situati sotto i nuclei. Il 17° giorno i nuclei, spinti da parte dai vacuoli, si trovano nella parte centrale della cellula. Il 18° giorno, i vacuoli si trovano nella parte apicale e i nuclei si trovano nella parte basale delle cellule, il glicogeno inizia a essere rilasciato nel lume delle ghiandole dalla secrezione apocrina. Le migliori condizioni per l'impianto si creano il 6-7° giorno dopo l'ovulazione, ad es. il 20-21° giorno del ciclo, quando l'attività secretoria delle ghiandole è massima.
Il 21° giorno del ciclo inizia la reazione decidua dello stroma endometriale. Le arterie spirali sono nettamente tortuose; successivamente, a causa di una diminuzione dell'edema dello stroma, sono chiaramente visibili. In primo luogo, compaiono le cellule deciduali, che gradualmente formano grappoli. Al 24° giorno del ciclo, questi accumuli formano manicotti eosinofili perivascolari. Il 25° giorno si formano isole di cellule deciduali. Entro il 26° giorno del ciclo, la reazione decidua diventa il numero di neutrofili che migrano lì dal sangue. L'infiltrazione neutrofila è sostituita dalla necrosi dello strato funzionale dell'endometrio.
Mestruazioni:
Se l'impianto non si verifica, le ghiandole cessano di produrre un segreto e iniziano i cambiamenti degenerativi nello strato funzionale dell'endometrio. La ragione immediata del suo rifiuto è un forte calo del contenuto di estradiolo e progesterone a causa dell'involuzione del corpo luteo. Nell'endometrio, il deflusso venoso diminuisce e si verifica vasodilatazione. Quindi si verifica un restringimento delle arterie, che porta a ischemia e danno tissutale e perdita funzionale dell'endometrio. Quindi si verifica il sanguinamento da frammenti di arteriole rimasti nello strato basale dell'endometrio. Le mestruazioni si fermano con il restringimento delle arterie, l'endometrio viene ripristinato. Pertanto, la cessazione del sanguinamento nei vasi dell'endometrio è diversa dall'emostasi in altre parti del corpo.
Di norma, l'emorragia si interrompe a causa dell'accumulo di piastrine e della deposizione di fibrina, che porta alla formazione di cicatrici. Nell'endometrio, la cicatrizzazione può portare alla perdita della sua attività funzionale (sindrome di Asherman). Per evitare queste conseguenze, è necessario un sistema alternativo di emostasi. La contrazione vascolare è un meccanismo per fermare l'emorragia nell'endometrio. Allo stesso tempo, le cicatrici sono ridotte al minimo dalla fibrinolisi, che distrugge i coaguli di sangue. Successivamente, il ripristino dell'endometrio e la formazione di nuovi vasi sanguigni (angiogenesi) porta al completamento del sanguinamento entro 5-7 giorni dall'inizio del ciclo mestruale.
L'effetto della sospensione di estrogeni e progesterone sulle mestruazioni è ben definito, ma il ruolo dei mediatori paracrini rimane poco chiaro. Vasocostrittori: la prostaglandina F2a, l'endoteliale-1 e il fattore di attivazione piastrinica (TAF) possono essere prodotti all'interno dell'endometrio e partecipare alla vasocostrizione. Contribuiscono anche all'inizio delle mestruazioni e a un ulteriore controllo su di esse. Questi mediatori possono essere regolati dall'azione di vasodilatatori come la prostaglandina E2, la prostaciclina, l'ossido nitrico, che sono prodotti dall'endometrio. La prostaglandina F2a ha un pronunciato effetto vasocostrittore, aumenta lo spasmo arterioso e l'ischemia endometriale, provoca contrazioni del miometrio, che, da un lato, riduce il flusso sanguigno e, dall'altro, aiuta a rimuovere l'endometrio rifiutato.
La riparazione endometriale comprende la rigenerazione ghiandolare e stromale e l'angiogenesi. Il fattore di crescita endoteliale vascolare (VEGF) e il fattore di crescita fibroplastico (FGF) si trovano nell'endometrio e sono forti agenti di angiogenesi. È stato riscontrato che la rigenerazione ghiandolare e stromale prodotta dagli estrogeni è migliorata sotto l'influenza dei fattori di crescita epidermica (EGF). Fattori di crescita come il fattore di crescita trasformante (TGF) e le interleuchine, in particolare l'interleuchina-1 (IL-1), hanno Grande importanza.
Una breve panoramica dei processi che si verificano nell'endometrio
Mestruazioni:
Il ruolo principale all'inizio delle mestruazioni è svolto dallo spasmo delle arteriole.
Lo strato funzionale dell'endometrio (superiore, che costituisce il 75% dello spessore) viene rifiutato.
Le mestruazioni si fermano a causa del vasospasmo e del ripristino dell'endometrio. La fibrinolisi previene la formazione di aderenze.
Fase proliferativa:
È caratterizzato dalla proliferazione delle ghiandole e dello stroma indotta dagli estrogeni.
fase secretoria:
È caratterizzato dalla secrezione di ghiandole indotta dal progesterone.
Nella fase secretoria tardiva viene indotta la decidualizzazione.
La decidualizzazione è un processo irreversibile. In assenza di gravidanza, l'apoptosi si verifica nell'endometrio, seguita dalla comparsa delle mestruazioni.
Quindi, il sistema riproduttivo è un supersistema, il cui stato funzionale è determinato dall'afferentazione inversa dei suoi sottosistemi costitutivi. Assegna: un lungo ciclo di feedback tra gli ormoni dell'ovaio e i nuclei dell'ipotalamo; tra gli ormoni ovarici e la ghiandola pituitaria; un breve anello tra l'ipofisi anteriore e l'ipotalamo; ultracorto tra RG-LH e neurociti (cellule nervose) dell'ipotalamo.
Il feedback di una donna sessualmente matura è sia negativo che positivo. Un esempio di associazione negativa è un aumento del rilascio di LH dalla ghiandola pituitaria anteriore in risposta a bassi livelli di estradiolo nella fase follicolare iniziale del ciclo. Un esempio di feedback positivo è il rilascio di LH e FSH in risposta al massimo ovulatoria di estradiolo nel sangue. Secondo il meccanismo del feedback negativo, la formazione di RG-LH aumenta con una diminuzione del livello di LH nelle cellule della ghiandola pituitaria anteriore.
Riepilogo
Il GnRH è sintetizzato dai neuroni del nucleo dell'infundibolo, quindi entra nel sistema portale della ghiandola pituitaria ed entra attraverso di esso nell'adenoipofisi. La secrezione di GnRH avviene impulsivamente.
La fase iniziale di sviluppo del gruppo follicolare primordiale è indipendente dall'FSH.
Quando il corpo luteo si evolve, la secrezione di progesterone e inibina diminuisce e i livelli di FSH aumentano.
L'FSH stimola la crescita e lo sviluppo di un gruppo di follicoli primordiali e la loro secrezione di estrogeni.
Gli estrogeni preparano l'utero all'impianto stimolando la proliferazione e la differenziazione dello strato funzionale dell'endometrio e, insieme all'FSH, favoriscono lo sviluppo dei follicoli.
Secondo la teoria delle due cellule della sintesi degli ormoni sessuali, l'LH stimola la sintesi degli androgeni nei tecociti, che vengono poi convertiti in estrogeni nelle cellule della granulosa sotto l'influenza dell'FSH.
Un aumento della concentrazione di estradiolo da un meccanismo di feedback negativo, un ciclo
che chiude nell'ipofisi e nell'ipotalamo, sopprime la secrezione di FSH.
Il follicolo che ovulerà in un dato ciclo mestruale è chiamato follicolo dominante. A differenza di altri follicoli che hanno iniziato a crescere, trasporta Di più Recettori dell'FSH e sintetizzano più estrogeni. Ciò gli consente di svilupparsi nonostante la diminuzione dei livelli di FSH.
Una stimolazione estrogenica sufficiente fornisce un picco di LH ovulatoria. A sua volta, provoca l'ovulazione, la formazione del corpo luteo e la secrezione di progesterone.
Il funzionamento del corpo luteo dipende dal livello di LH. Con la sua diminuzione, il corpo luteo subisce un'involuzione. Questo di solito si verifica il 12-16° giorno dopo l'ovulazione.
Se è avvenuta la fecondazione, l'esistenza del corpo luteo è supportata dalla gonadotropina corionica. Il corpo luteo continua a secernere progesterone, necessario per mantenere la gravidanza nelle prime fasi.
La regolazione della funzione degli organi genitali è svolta da un complesso sistema neuroumorale autoregolante.
Il ruolo principale in esso è svolto dal cervello e prima di tutto dalla corteccia cerebrale. Il centro del sistema è il complesso ipotalamo - ghiandola pituitaria - ovaie.
Il sistema nervoso autonomo, le prostaglandine, le sostanze biologicamente attive della ghiandola pineale, gli ormoni tiroidei, surrenali e pancreatici influenzano.
Ipotalamo- il più alto centro di regolazione delle funzioni endocrine del corpo. Nell'ipotalamo si trovano i neuroni che percepiscono tutti i cambiamenti che si verificano nel corpo. Questo riceve informazioni dai centri che regolano l'attività dei sistemi respiratorio e cardiovascolare.
Nell'ipotalamo ci sono centri della sete, della fame, centri che regolano le funzioni sessuali, le emozioni e il comportamento umano, il sonno e la veglia, la temperatura corporea e le reazioni vegetative.
L'ipotalamo secerne fattori di rilascio- sostanze che regolano l'attività di un'altra importante ghiandola endocrina - la ghiandola pituitaria.
pituitario situato nelle profondità della sella turca dell'osso sfenoide del cranio, ha due lobi: anteriore e posteriore. Sotto l'influenza dei fattori di rilascio dell'ipotalamo, la ghiandola pituitaria produce ormoni che regolano il funzionamento delle ghiandole endocrine: la tiroide, le ghiandole surrenali e le gonadi.
Le ovaie funzionano sotto l'influenza ormoni gonadotropi ipofisario: follicolo-stimolante (FSH) e luteinizzante (LH), nonché prolattina (Prl). La formazione di estrogeni, progesterone e androgeni nelle ovaie dipende dal loro livello. La prolattina supporta la funzione del corpo luteo e influisce anche sulla secrezione di latte nel periodo postpartum.
Corpo pineale (ghiandola pineale)- una ghiandola spaiata situata nel mesencefalo, sopra il cervelletto. È chiamato "l'orologio biologico" del corpo.
Il corpo pineale ha un effetto inibitorio sulla funzione dell'ipotalamo, che è di grande importanza nelle malattie ginecologiche, nella gravidanza, nello sviluppo del travaglio, nell'allattamento.
ghiandole surrenali producono ormoni che regolano il metabolismo dei carboidrati, delle proteine e dei minerali, androgeni, estrogeni e ormoni dello stress: adrenalina, norepinefrina, dopamina. Questi ultimi causano vasocostrizione, contrazione uterina, hanno un effetto inibitorio su funzione ormonale ovaie.
Prostaglandine- Si tratta di sostanze che sono vicine nella loro azione agli ormoni classici, ma sono sintetizzate nelle cellule di vari tessuti del corpo. Le prostaglandine hanno un effetto pronunciato sul sistema endocrino, in particolare sulla ghiandola pituitaria, sulle ovaie, riducono l'attività funzionale del corpo luteo.
Le prostaglandine migliorano la contrattilità dell'utero, la loro introduzione interrompe la gravidanza di qualsiasi periodo, svolgono un ruolo importante nell'inizio del parto spontaneo, nella preeclampsia tardiva, nella debolezza dell'attività lavorativa.
Quindi il regolamento Il ciclo mestruale avviene in un sistema graduale: la corteccia cerebrale - l'ipotalamo - la ghiandola pituitaria - le ovaie - gli organi bersaglio (utero, cervice, ghiandole mammarie, ecc.). Nelle cellule dell'ipotalamo vengono prodotti gli ormoni di rilascio, sotto l'azione dei quali vengono prodotti FSH e LH nella ghiandola pituitaria.
Gli ormoni gonadotropici secreti dalla ghiandola pituitaria causano la secrezione di estrogeni e progesterone nell'ovaio in modo feed-forward. L'ovulazione avviene sotto l'influenza di FSH e LH.
Quando si raggiunge un livello significativo di ormoni ovarici, questi ultimi determinano un effetto inibitorio sulla secrezione degli ormoni gonadotropici secondo il principio del feedback. Sullo sfondo di un declino livello generale si verificano ormoni dell'ipotalamo, della ghiandola pituitaria, delle ovaie, delle mestruazioni.
M ciclo mestrualeè un complesso processo biologico in cui sono coinvolti molti sistemi e organi del corpo, mentre l'attività dell'utero è la fase finale di questi processi.


